Одомашнивание овец.
Ученые считают, что одомашнивать овец начали примерно 8 — 12 тысяч лет назад на обширной территории от Каспийского моря до Гималаев. Приручали этих животных жители Южной Европы, Северной Америки и Северной Африки.
Пойманных козлят и ягнят охотники не убивали, а сохраняли про запас возле поселений. Как только овцы и козы начали размножаться в неволе и их численность стала возрастать, им потребовались пастбища. Это вызвало необходимость частого перегона стад, и люди стали вести кочевой образ жизни.
На Среднем Востоке жизнь пастухов находилась в полной зависимости от овец, поэтому животные жили среди людей на правах любимых детей. Ягненок здесь отождествлялся с младенцем, их крик одинаково «брал за сердце», и им уделялось равное внимание. В течение столетий овец доили главным образом в странах Средиземноморья и Среднего Востока, где засуха делала немыслимой любую другую отрасль животноводства.
 Полагают, что они произошли от
диких видов, обитавших в Малой Азии,
Персии и соседних странах. В Библии козы
упоминаются неоднократно. На древнейших
гробницах фараонов имеются барельефы
с изображениями этих животных, общипывающих
листву на верхней части деревьев..
Полагают, что они произошли от
диких видов, обитавших в Малой Азии,
Персии и соседних странах. В Библии козы
упоминаются неоднократно. На древнейших
гробницах фараонов имеются барельефы
с изображениями этих животных, общипывающих
листву на верхней части деревьев..В Норвегии овцы занимает особое, почетное положение в обществе. Легенда рассказывает, что норвежский король Олаф II однажды спас раненую овцу. Животное доставили во дворец, вылечили и потом отпустили восвояси. В знак благодарности она каждую ночь приносила спасителю редкие целебные растения. И по сей день подарки детям в этой стране раздает овца, которую угощают овсяной соломой. Запряженная в саночки, она доставляет большое удовольствие детворе.
Еще
5 — 6 тысяч лет назад горожанки Египта и
Вавилона считали самой модной одеждой
костюм из тонкой шерсти. Это очень
стимулировало развитие тонкосуконной
промышленности, а пастухам пришлось
незамедлительно приступить к выведению
новых пород тонкорунных овец с длинным
и очень тонким подшерстком.
После
падения Римской империи
тонкорунное овцеводство почти
исчезло. Спрос на тонкую шерсть
возобновился только в эпоху Возрождения.
Обладательницей тонкорунных овец
оказалась одна Испания, которая не
упустила возможности использовать свое
положение. Под страхом смертной казни
запрещалось вывозить из страны живых
тонкорунных овец — мериносов. В то время
в Испании было широко распространено
отгонное овцеводство. В летнюю пору
овец пасли на прохладных горных пастбищах,
а с наступлением зимы перегоняли в
долины. Считают, что слово «меринос»
испанского происхождения и означает
«странствующий», «бродячий».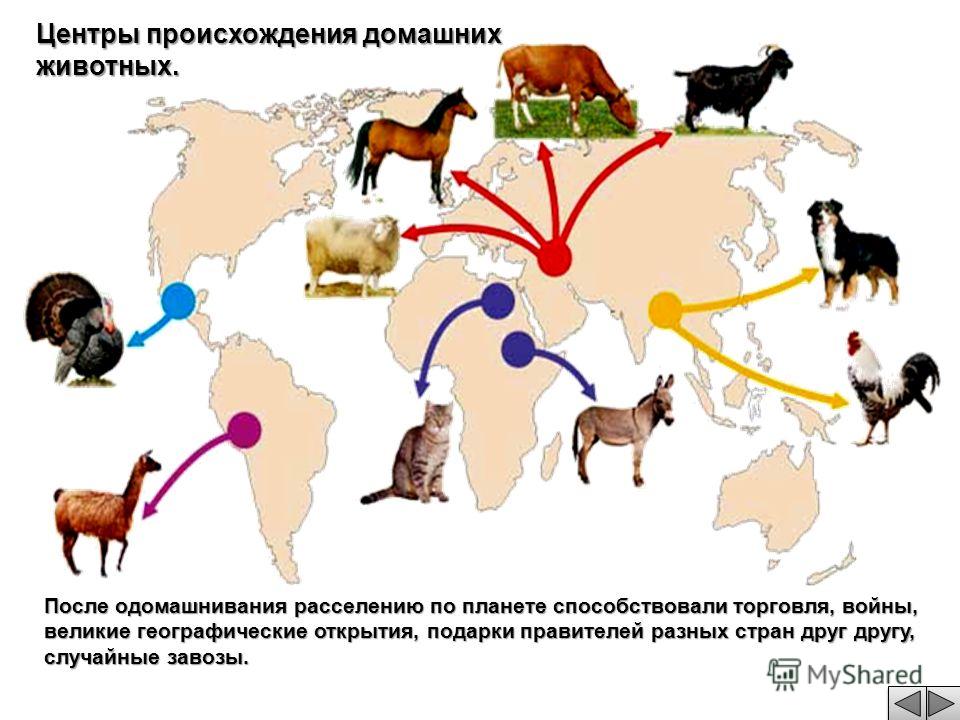
В старых арабских медицинских книгах упоминается, что о бодрящих свойствах кофейных зерен людям «поведали» овцы. Монах-пастух одного из монастырей в Эфиопии подметил, что в те дни, когда овцы паслись на склонах горы, поросших кофейными кустами и деревьями, они становились особенно оживленными, игривыми. Возбуждение не проходило и ночью в загонах. Этому пастуху, как и прочим монахам, после тяжелого трудового дня приходилось принимать участие в ночных бдениях. Чтобы побороть неодолимую сонливость, он попробовал есть плоды кофе. И сон отступил! Вскоре его примеру последовали другие монахи, а затем этот опыт распространился за пределами монастыря.
Известны
и современные «открытия» животных.
Так, источник минеральной воды в Уральской
области открыли овцы. Чабаны неоднократно
замечали, что стоит лишь пригнать отару
в небольшую ложбину, как тут же овцы
устремляются к маленькому роднику на
склоне сопки. Как было затем установлено,
в воде источника содержатся биологически
активные компоненты и другие вещества,
имеющие целебные свойства.
Основные районы разведения овец: Казахстан, Средняя Азия и Закавказье, Северный Кавказ, Украина, Поволжье, Урал, Западная и Восточная Сибирь.
Были даже попытки провести классификацию домашних пород, связав их происхождение от той или иной дикой формы: одних — от европейского муфлона, других — от архара, третьих — от аргали. Однако такие сближения оказывались натянутыми, так как при переселении племен различные расы одомашненных овец смешивались между собой, да по существу эти сближения были и ненужными, раз все их родоначальные формы могут быть отнесены к одному зоологическому виду.
Видовое единство домашних овец и диких баранов выражается и в получении от них вполне плодовитых помесей.
В
общем история одомашнивания овцы
несколько напоминает историю одомашнивания
собаки. По-видимому, этот процесс
совершался независимо в различных
областях, где обитали разные подвиды
диких баранов, а позднее, при переселениях
человеческих племен, восточные овцы
смешались с одомашненными европейскими
расами. В более новые времена, в связи
с захватом европейцами заморских
колоний, овцы распространились человеком
далеко за пределы их первоначальной
родины, и в настоящее время овцеводство
процветает в различных областях Австралии
и Аргентины.
В более новые времена, в связи
с захватом европейцами заморских
колоний, овцы распространились человеком
далеко за пределы их первоначальной
родины, и в настоящее время овцеводство
процветает в различных областях Австралии
и Аргентины.
Одомашненная овца оказалась для наших предков очень ценным приобретением. Доставляя молоко, мясо, сало, шерсть и овчину, овца одевала, и кормила своего хозяина и даже снабжала его материалом для его легкого переносного жилища.
Антропологи разобрались в одомашнивании овец и коз в Ашиклы-Хююке
Антропологи исследовали кости коз и овец, обнаруженные в ходе раскопок неолитического поселения Ашиклы-Хююк (около 8400–7350 годов до нашей эры). Ученые выяснили, что на раннем этапе занятия этой стоянки местные жители начали отлавливать диких ягнят и козлят, после чего выращивали их до возраста 6–12 месяцев. Постепенно стали появляться свидетельства того, что животные начали доживать до полового созревания и давать потомство. На поздних этапах существования поселения уже видны черты крупного скотоводческого хозяйства, когда многие овцы и козы доживали до зрелого возраста. Результаты исследования опубликованы в журнале Proceedings of the National Academy of Sciences.
Результаты исследования опубликованы в журнале Proceedings of the National Academy of Sciences.
Овцы и козы оказались одними из древнейших животных, одомашненных человеком. Палеозоологические данные указывают на то, что процесс доместикации этих животных начался в обширном регионе, простирающемся от Восточной Анатолии до Ирана и Ирака, где люди начали отлавливать и выращивать диких парнокопытных млекопитающих – азиатского муфлона (Ovis gmelini) и безоарового козла (Capra aegagrus). Вместе с пришлым неолитическим населением около 6000 года до нашей эры культура разведения этих животных распространилась на Европу и Средиземноморье, а около 5000 года до нашей эры – на Северную Африку. В конце IV тысячелетия до нашей эры первые скотоводы, разводившие коз и овец, прибыли во Внутреннюю Азию.
В 25 километрах от современного турецкого города Аксарай расположен курган из вулканического туфа, в котором археологи обнаружили доисторическое поселение Ашиклы-Хююк, возникшее в середине IX тысячелетия до нашей эры. Оно представляет собой одну из самых ранних стоянок докерамического неолита в Центральной Анатолии, где археологи обнаружили свидетельства постепенного перехода от охоты к скотоводству, который растянулся примерно на 1000 лет.
Оно представляет собой одну из самых ранних стоянок докерамического неолита в Центральной Анатолии, где археологи обнаружили свидетельства постепенного перехода от охоты к скотоводству, который растянулся примерно на 1000 лет.
Мэри Стайнер (Mary Stiner) из Аризонского университета совместно с учеными из Нидерландов, США и Турции исследовала процесс одомашнивания коз и овец в поселении Ашиклы-Хююк, где была собрана богатая коллекция фаунистических материалов, относящихся к тысячелетнему периоду (около 8400–7350 годов до нашей эры). Антропологи отметили, что этот памятник находится внутри ареалов муфлона (Ovis gmelinii) и безоарового козла (Capra aegagrus).
Ученые обнаружили, что в самом древнем культурном слое находились кости крупной и мелкой дикой добычи, причем, судя по количеству останков, козы и овцы первоначально были второстепенным источником мяса (около 27 процентов). Однако постепенно их доля достигла 81 процента. Антропологи отметили, что на протяжении всех стадий оккупации стоянки овцам отдавалось предпочтение перед козами. Кроме того, анализ навоза показал, что этих животных с самого начала существования поселения содержали в неволе на территории памятника.
Кроме того, анализ навоза показал, что этих животных с самого начала существования поселения содержали в неволе на территории памятника.
Результаты исследования продемонстрировали, что в течение тысячи лет последовательно сменяли друг друга три формы взаимодействия между человеком и козлиными. На ранней стадии существования поселения люди отлавливали живых ягнят и козлят в дикой природе вскоре после их рождения, после чего в течение нескольких месяцев выращивали их в поселении (как правило, животных убивали в возрасте 6–12 месяцев). Такая стратегия, видимо, объясняется хранением «живого» мяса вплоть до возможного сезона голода или до важного общественного мероприятия. Похожие практики известны и в некоторых современных культурах собирателей и земледельцев.
Однако ученые заметили, что уже в самых древних культурных слоях постепенно увеличивалось количество костей недоношенных или новорожденных животных — свидетельства того, что козы и овцы начали размножаться в неволе. В более позднее время антропологи нашли признаки крупного скотоводческого хозяйства, когда многим взрослым овцам и козам обоих полов позволяли доживать до зрелого возраста. Кроме того, в это время наблюдалось снижение числа недоношенных особей, а также улучшение состояния здоровья самих животных.
Кроме того, в это время наблюдалось снижение числа недоношенных особей, а также улучшение состояния здоровья самих животных.
Антропологи также обнаружили, что за тысячелетний период существования поселения Ашиклы-Хююк ни у коз, ни у баранов не произошло заметного уменьшения размеров тела. По всей видимости, это объясняется тем, что на протяжении всего периода местные жители продолжали разбавлять стада небольшим количеством отловленных диких ягнят и козлят. Изотопные анализы показали, что, несмотря на примерно десятикратный рост количества населения в Ашиклы-Хююке за тысячелетие, потребление человеком животного белка оставалось примерно на одном уровне, что указывает на существенный рост поголовья скота. Это также подтверждается анализами солей мочи в культурных слоях.
Ранее на N + 1 рассказывали о других исследованиях, посвященных доместикации диких животных. Так, археологи обнаружили древнейшие свидетельства разведения буйволов на Кавказе. Кроме того, генетики выяснили, что кубанские туры передали домашним козам защиту кишечника.
Михаил Подрезов
Нашли опечатку? Выделите фрагмент и нажмите Ctrl+Enter.
РАСКРЫТИЕ ИСТОРИИ ДОМЕСТИРОВАНИЯ ОВЕЦ С ИСПОЛЬЗОВАНИЕМ ИНТЕГРАЦИИ РЕТРОВИРУСОВ
- Список журналов
- Рукописи авторов ЧВК Европы
- PMC3145132
Наука. Авторская рукопись; доступно в PMC 2011 28 июля.
Опубликовано в окончательной редакции как:
Наука. 2009 г., 24 апреля; 324 (5926): 532–536.
doi: 10.1126/science.1170587
PMCID: PMC3145132
EMSID: UKMS35940
PMID: 193
, 1, 2 , 3 , 1 , 3 , 4 , 5 , 1 , 6 , 6 , 1 , 2 , 2 , 7 , 8 , 9 , 10 , 11 , 12 , 13 , 14 , 15 , 16 , 17 , 18 , 19 , 20 , 21 , 22 , 23 , 24 , 25 , 26 , 27 и 1, *
Авторская информация о сообщении об оплате и лицензии
- 903 представляло собой решающий шаг в истории человечества.
 Используя в качестве генетических маркеров эндогенные ретровирусы, мы обнаружили, что овцы дифференцировались на основе своего «ретротипа» и морфологических признаков, рассредоточенных по Евразии и Африке посредством отдельных миграционных эпизодов. Реликты первых миграций включают муфлонов, а также породы, ранее считавшиеся «примитивными» на основании их морфологии, такие как оркнейские, соайские и нордические короткохвостые овцы, в настоящее время обитающие на периферии северо-западной Европы. Более поздний миграционный период с участием овец с улучшенными производственными качествами сформировал подавляющее большинство современных пород. Возможность отличить генетически примитивных овец от более современных пород дает ценную информацию об истории одомашнивания овец.
Используя в качестве генетических маркеров эндогенные ретровирусы, мы обнаружили, что овцы дифференцировались на основе своего «ретротипа» и морфологических признаков, рассредоточенных по Евразии и Африке посредством отдельных миграционных эпизодов. Реликты первых миграций включают муфлонов, а также породы, ранее считавшиеся «примитивными» на основании их морфологии, такие как оркнейские, соайские и нордические короткохвостые овцы, в настоящее время обитающие на периферии северо-западной Европы. Более поздний миграционный период с участием овец с улучшенными производственными качествами сформировал подавляющее большинство современных пород. Возможность отличить генетически примитивных овец от более современных пород дает ценную информацию об истории одомашнивания овец. Первые сельскохозяйственные системы, основанные на выращивании зерновых, бобовых и разведении домашнего скота, появились в Юго-Западной Азии примерно за 11 000 лет до настоящего времени (YBP) (1, 2). К 6000 лет назад агро-скотоводство, введенное неолитической сельскохозяйственной революцией, стало основной системой производства продуктов питания во всей доисторической Европе, от севера Средиземноморья до Британии, Ирландии и Скандинавии (3), на юге до Северной Африки (4) и с востока до запада и Центральная Азия (5).
Овцы и козы были первыми видами домашнего скота, которые были одомашнены (6). Множественные события одомашнивания, как это следует из множества митохондриальных линий, привели к появлению домашних овец и аналогичных других домашних видов (7-10). Первоначально овец разводили в основном на мясо, но в течение пятого тысячелетия до нашей эры в Юго-Западной Азии и четвертого тысячелетия до нашей эры в Европе стала очевидной специализация на «вторичных» продуктах, таких как шерсть. Овцы, отобранные для производства вторичных продуктов, по-видимому, заменили более примитивное домашнее население. Вопрос о том, произошла ли специализация на производстве вторичных продуктов сначала в Юго-Западной Азии или во всей Европе, точно не известен из-за отсутствия точных археологических свидетельств начала производства шерсти (6, 11, 12).
В этом исследовании мы использовали семейство эндогенных ретровирусов (ERV) в качестве генетических маркеров для изучения истории домашних овец. ERV возникают в результате стабильной интеграции генома ретровируса («провируса») в зародышевую линию хозяина (13) и передаются вертикально из поколения в поколение по менделевскому типу. Геном овцы содержит не менее 27 копий эндогенных ретровирусов, связанных с экзогенным и патогенным ретровирусом овец Яагсиекте (enJSRV) (14-16). Большинство локусов enJSRVs фиксируются у домашних овец, но некоторые по-разному распределяются между породами и особями (т.е. они инсерционно полиморфны) (14). enJSRV можно использовать в качестве высокоинформативных генетических маркеров, поскольку присутствие каждого эндогенного ретровируса в геноме хозяина является результатом единственного события интеграции у одного животного и является необратимым, поэтому популяции, имеющие один и тот же провирус в одном и том же месте генома, необратимы.0013 де-факто
Геном овцы содержит не менее 27 копий эндогенных ретровирусов, связанных с экзогенным и патогенным ретровирусом овец Яагсиекте (enJSRV) (14-16). Большинство локусов enJSRVs фиксируются у домашних овец, но некоторые по-разному распределяются между породами и особями (т.е. они инсерционно полиморфны) (14). enJSRV можно использовать в качестве высокоинформативных генетических маркеров, поскольку присутствие каждого эндогенного ретровируса в геноме хозяина является результатом единственного события интеграции у одного животного и является необратимым, поэтому популяции, имеющие один и тот же провирус в одном и том же месте генома, необратимы.0013 де-факто
Мы проанализировали образцы геномной ДНК, собранные у 1362 животных, принадлежащих к 133 породам домашних овец ( Ovis orientalis aries , обычно обозначаемых как Ovis aries ) и ближайших диких родственников (см. ниже), разделенных на 65 групп, образованных одним или больше пород, имеющих общее географическое положение и/или связи для разведения (таблица S1) (17). Исследуемые образцы также включали уриальных овец ( Ovis vignei ), средиземноморских и азиатских муфлонов ().0013 Ovis orientalis musimon, Ovis orientalis ophion и Ovis orientalis orientalis ). Подавляющее большинство изученных нами пород являются локальными, исторически связанными с конкретными географическими районами и не подверженными интенсивным программам разведения товарных стад.
Исследуемые образцы также включали уриальных овец ( Ovis vignei ), средиземноморских и азиатских муфлонов ().0013 Ovis orientalis musimon, Ovis orientalis ophion и Ovis orientalis orientalis ). Подавляющее большинство изученных нами пород являются локальными, исторически связанными с конкретными географическими районами и не подверженными интенсивным программам разведения товарных стад.
Образцы тестировали на наличие или отсутствие шести независимо наследуемых инсерционно полиморфных enJSRV (enJSRV-18, enJSRV-7, enJSRV-8, enJSRV-15, enJSRV-16 и enJS5F16) с помощью полимеразной цепной реакции (ПЦР) с использованием двух наборы праймеров, которые амплифицируют соответственно 5′- и 3′-концевые повторы (LTR) каждого провируса (включая фланкирующие последовательности геномной ДНК хозяина), как описано ранее (14, 17). Провирус enJSRV-18 имел самую высокую частоту в нашем наборе данных (85%), enJSRV-7 и enJS5F16 были обнаружены в 27% и 30% образцов соответственно, тогда как enJSRV-15, enJSRV-16 и enJSRV-8 были обнаружены в 27% и 30% образцов соответственно. присутствует только в 3-5% образцов ().
присутствует только в 3-5% образцов ().
Открыть в отдельном окне
Всемирное распространение инсерционно-полиморфных enJSRV. Распределение инсерционно-полиморфных локусов enJSRV проанализировано в этом исследовании в 65 популяциях овец, представляющих местные породы Старого Света. (A) Частоты каждого локуса enJSRV в каждой популяции представлены вертикальной чертой и расположены в порядке убывания. Частоты вставок были получены с использованием программного обеспечения Arlequin 3.11 (27), обрабатывающего отсутствие специфического провируса enJSRV как рецессивный аллель. (B) Местоположение отобранных популяций овец. (C-F) . Карты интерполяции, отображающие пространственное распределение предполагаемых частот enJSRV. Географические вариации были визуализированы с помощью «Spatial Analyst Extension» программного обеспечения ArcView GIS 3.2 (ESRI, Редлендс, Калифорния, США; http://www.esri.com). Интерполированные значения карты были рассчитаны с использованием обратного взвешивания по расстоянию с 12 ближайшими соседями и степенью двойки, а поверхности интерполяции были разделены на 13 классов с более высокими частотами вставки, обозначенными самой темной штриховкой. В качестве географических координат для каждой популяции использовалась центральная точка области отбора проб (таблица S1).
В качестве географических координат для каждой популяции использовалась центральная точка области отбора проб (таблица S1).
Мы сделали вывод о распределении инсерционно-полиморфных локусов enJSRV у самых первых домашних овец, определив их встречаемость у уриальных овец и средиземноморских/азиатских муфлонов, а затем проверив молекулярные сигнатуры, указывающие на возраст провируса. Предполагаемое расхождение между уриалом (одним из ближайших ныне живущих родственников домашней овцы) и домашней овцой составляет примерно 800 000 YBP (18). Следовательно, любой провирус, общий для этих двух видов, предшествует процессу одомашнивания. То же верно и для азиатского муфлона, который считается прямым предком домашней овцы (19).-21), в то время как близкородственный средиземноморский муфлон считается остатком первой одомашненной овцы, повторно приспособленной к дикой жизни (19, 22, 23). Несмотря на широкое распространение в протестированных образцах, enJSRV-18 отсутствовал у уриальных овец (n=5), средиземноморских муфлонов (n=17) и азиатских муфлонов (n=15). Напротив, относительно более редкий enJSRV-7 был обнаружен у трех из пяти уриальных овец, у большинства (86%) азиатских муфлонов и у всех средиземноморских муфлонов. Эти данные предполагают, что интеграция enJSRV-7 в зародышевую линию хозяина предшествовала интеграции enJSRV-18. Различия между проксимальными (5′) и дистальными (3′) длинными концевыми повторами (LTR) enJSRV-7 подтверждают его предшественник. Расхождение между 5′- и 3′-LTR эндогенного ретровируса дает оценку «возраста» эндогенного провируса, потому что при заражении ретровирусы обратно транскрибируют свой геном из РНК в ДНК, и во время этого процесса они дублируют геномные концы, давая начало на два одинаковых LTR. Проксимальные и дистальные LTR эндогенного ретровируса должны быть идентичны при интеграции, но могут расходиться с течением времени с той же скоростью, что и некодирующие последовательности (~ 2,3–5 × 10 -9 замен на сайт в год). enJSRV-7, по-видимому, является самым старым провирусом в наших образцах, поскольку он демонстрирует пять нуклеотидных замен между 5′ и 3′ LTR (длиной 445 нуклеотидов), в то время как все другие инсерционно-полиморфные провирусы (включая enJSRV-18) обладают идентичными LTR.
Напротив, относительно более редкий enJSRV-7 был обнаружен у трех из пяти уриальных овец, у большинства (86%) азиатских муфлонов и у всех средиземноморских муфлонов. Эти данные предполагают, что интеграция enJSRV-7 в зародышевую линию хозяина предшествовала интеграции enJSRV-18. Различия между проксимальными (5′) и дистальными (3′) длинными концевыми повторами (LTR) enJSRV-7 подтверждают его предшественник. Расхождение между 5′- и 3′-LTR эндогенного ретровируса дает оценку «возраста» эндогенного провируса, потому что при заражении ретровирусы обратно транскрибируют свой геном из РНК в ДНК, и во время этого процесса они дублируют геномные концы, давая начало на два одинаковых LTR. Проксимальные и дистальные LTR эндогенного ретровируса должны быть идентичны при интеграции, но могут расходиться с течением времени с той же скоростью, что и некодирующие последовательности (~ 2,3–5 × 10 -9 замен на сайт в год). enJSRV-7, по-видимому, является самым старым провирусом в наших образцах, поскольку он демонстрирует пять нуклеотидных замен между 5′ и 3′ LTR (длиной 445 нуклеотидов), в то время как все другие инсерционно-полиморфные провирусы (включая enJSRV-18) обладают идентичными LTR. Эти данные свидетельствуют о том, что популяции, происходящие от самых первых домашних овец, не несли ни одного из инсерционно-полиморфных enJSRV, использованных в этом исследовании, или несли enJSRV-7.
Эти данные свидетельствуют о том, что популяции, происходящие от самых первых домашних овец, не несли ни одного из инсерционно-полиморфных enJSRV, использованных в этом исследовании, или несли enJSRV-7.
Географическая изменчивость всех локусов enJSRV была визуализирована путем построения карт интерполяции на основе их значений частоты вставки (). Самая высокая частота enJSRV-7 была обнаружена у средиземноморских муфлонов и у овец сой, населяющих в настоящее время остров Сент-Килда у северо-запада Шотландии. enJSRV-18 был равномерно распределен на очень высоких частотах по всему Старому Свету. Низкие частоты enJSRV-18 наблюдались на островах, населенных средиземноморскими муфлонами, и в периферийных районах СЗ Европы. Два провируса enJSRV, enJS5F16 и enJSRV-8, показали схожий географический паттерн с высокой частотой на Британских островах и в Скандинавии. Менее распространенные enJSRV-15 и enJSRV-16 имели менее очевидные географические закономерности (рис. S1).
Затем мы проанализировали комбинацию инсерционно-полиморфных enJSRV (которую мы называем «ретротипом») в каждой из проанализированных популяций (). Ретротип R2 (представляющий наличие только enJSRV-18) был преобладающим ретротипом в большинстве протестированных популяций. Интересно, что ретротип R4, указывающий на совместное присутствие enJSRV-18 и enJSRV-7 (), был еще одним распространенным ретротипом в области, соответствующей исторической Финикии, и в Южной Европе, предполагая, что морская торговля и колонизация оказали большое влияние на движение овец в Европе. Средиземноморья, что подтверждается исследованиями с использованием вариаций митохондриальной ДНК овец (24, 25). Дополнительные вставки enJSRV объясняют более сложные ретротипы популяций в Северной Европе (см. также вспомогательный онлайн-текст). Популяции овец в Африке, Пакистане и Китае демонстрировали аналогичный гомогенный ретротип R2, характерный для популяций в Юго-Западной Азии, что предполагает прямые миграционные связи домашних овец между этими районами. Большинство популяций из Скандинавии демонстрировали сходные ретротипы с населением Исландии и Фарерских островов, подтверждая исторически зарегистрированные перемещения норвежских поселенцев в конце первого тысячелетия нашей эры (26).
Ретротип R2 (представляющий наличие только enJSRV-18) был преобладающим ретротипом в большинстве протестированных популяций. Интересно, что ретротип R4, указывающий на совместное присутствие enJSRV-18 и enJSRV-7 (), был еще одним распространенным ретротипом в области, соответствующей исторической Финикии, и в Южной Европе, предполагая, что морская торговля и колонизация оказали большое влияние на движение овец в Европе. Средиземноморья, что подтверждается исследованиями с использованием вариаций митохондриальной ДНК овец (24, 25). Дополнительные вставки enJSRV объясняют более сложные ретротипы популяций в Северной Европе (см. также вспомогательный онлайн-текст). Популяции овец в Африке, Пакистане и Китае демонстрировали аналогичный гомогенный ретротип R2, характерный для популяций в Юго-Западной Азии, что предполагает прямые миграционные связи домашних овец между этими районами. Большинство популяций из Скандинавии демонстрировали сходные ретротипы с населением Исландии и Фарерских островов, подтверждая исторически зарегистрированные перемещения норвежских поселенцев в конце первого тысячелетия нашей эры (26). Чтобы визуализировать генетическое родство протестированных популяций, мы проанализировали данные, используя два разных подхода: график многомерного масштабирования (MDS), полученный из межпопуляционной матрицы несмещенных генетических расстояний Нея, и анализ главных компонентов (PCA), вычисленный из матрицы корреляции между вставками enJSRV. частоты.
Чтобы визуализировать генетическое родство протестированных популяций, мы проанализировали данные, используя два разных подхода: график многомерного масштабирования (MDS), полученный из межпопуляционной матрицы несмещенных генетических расстояний Нея, и анализ главных компонентов (PCA), вычисленный из матрицы корреляции между вставками enJSRV. частоты.
Открыть в отдельном окне
Комбинация провирусов enJSRV (ретротипов) у домашних овец. Круговые диаграммы на рисунке представляют частоту каждого ретротипа в 65 протестированных популяциях. Каждой протестированной овце был присвоен ретротип на основе комбинации инсерционно-полиморфных провирусов enJSRV, присутствующих в их геноме. Ретротипы определяли с R0 по R14 следующим образом: RO = нет инсерционно полиморфных enJSRV ; R1 = enJSRV-7 ; R2 = enJSRV-18 ; R3 = enJS5F16 ; R4 = enJSRV-7 + enJSRV-18 ; R5 = enJSRV-7 + enJS5F16 ; R6 = enJSRV-18 + enJS5F16 ; R7 = enJSRV-7 + enJSRV-18+ enJS5F16 ; R8 = enJSRV-8 ; R9 = enJS5F16 + enJSRV-8 ; R10 = enJSRV-7 + enJS5F16 + enJSRV-8 ; R11 = enJSRV-18 + enJSRV-8 ; R12 = enJSRV-18 + enJS5F16 + enJSRV-8 ; R13 = enJSRV-7 + enJSRV-18 + enJSRV-8 ; R14 = enJSRV-7 + enJSRV-18 + enJS5F16 + enJSRV-8. Каждый ретротип представлен своим цветом (и узором), как показано на рисунке. Цифры рядом с каждой круговой диаграммой обозначают каждую из 65 протестированных популяций, как указано в таблице S1. Обратите внимание, что большинство популяций в Юго-Западной Азии, Центральной Азии, Южной Европе и Африке обладают R2 (т.е. наличием только enJSRV-18, показано зеленым цветом) в качестве преобладающего ретротипа. Вокруг Средиземноморского бассейна также высока доля R4, обусловленная современным присутствием enJSRV-7 и enJSRV-18 (показаны желтым цветом). Примитивные породы характеризуются высокой долей животных с R0 (отсутствие инсерционно-полиморфных провирусов, показано белым) или R1 (присутствие только enJSRV-7, показано красным). «Скандинавский» ретротип R3 (показан синим цветом) характеризовался низкой частотой enJSRV-18 и высокой частотой enJS5F16; Северные популяции также имели относительно высокую частоту овец, у которых не было протестировано ни одного из инсерционно-полиморфных провирусов.
Каждый ретротип представлен своим цветом (и узором), как показано на рисунке. Цифры рядом с каждой круговой диаграммой обозначают каждую из 65 протестированных популяций, как указано в таблице S1. Обратите внимание, что большинство популяций в Юго-Западной Азии, Центральной Азии, Южной Европе и Африке обладают R2 (т.е. наличием только enJSRV-18, показано зеленым цветом) в качестве преобладающего ретротипа. Вокруг Средиземноморского бассейна также высока доля R4, обусловленная современным присутствием enJSRV-7 и enJSRV-18 (показаны желтым цветом). Примитивные породы характеризуются высокой долей животных с R0 (отсутствие инсерционно-полиморфных провирусов, показано белым) или R1 (присутствие только enJSRV-7, показано красным). «Скандинавский» ретротип R3 (показан синим цветом) характеризовался низкой частотой enJSRV-18 и высокой частотой enJS5F16; Северные популяции также имели относительно высокую частоту овец, у которых не было протестировано ни одного из инсерционно-полиморфных провирусов.
Анализ MDS выявил заметное разделение (особенно очевидное в первом измерении) между подавляющим большинством домашних пород и внешней группой, состоящей из муфлонов, соайских овец, гебридских, оркнейских овец, исландских и нордических пород (). Сходные результаты были получены методом PCA (). В совокупности полученные нами данные показывают, что реликты первых миграций все еще присутствуют у муфлонов Сардинии, Корсики и Кипра, а также у пород в периферийных районах Северной Европы. На основе своих ретротипов эти примитивные популяции характеризуются отсутствием enJSRV-18 (зафиксирован у большинства современных пород) и либо наличием enJSRV-7 с высокой частотой, либо отсутствием инсерционно-полиморфных enJSRV (включая enJSRV-7). ). Напротив, ретротипы подавляющего большинства пород овец группируются вместе и характеризуются высокой частотой или фиксацией enJSRV-18.
Открыть в отдельном окне
Генетические дистанции между популяциями овец на основе частот вставки enJSRV. (A) Многомерный (MDS) масштабный график, рассчитанный по матрице несмещенных генетических расстояний Нея (программное обеспечение TFPGA 1.3) (28). Во всех анализах учитывался доминантный характер enJSRV как генетических маркеров. Матрица межпопуляционных расстояний была обобщена в двух измерениях с использованием анализа MDS, реализованного STATISTICA ‘9.9 программный пакет (StatSoft Inc., Талса, Оклахома, США). Каждый треугольник представляет одну из 65 протестированных популяций. Обратите внимание, что на графике названы только те популяции, которые находятся за пределами основного кластера (обведены квадратом с пунктирной линией и включают подавляющее большинство пород из Африки, Азии и Европы). (B) Трехмерный график, суммирующий данные, полученные с помощью анализа главных компонентов (PCA) инсерционно-полиморфных провирусов enJSRV в 65 популяциях овец, протестированных с использованием Proc Factor статистического пакета SAS/STAT® (SAS Institute Inc, Cary NC ) по рекомендациям Кавалли-Сфорца и др.
(A) Многомерный (MDS) масштабный график, рассчитанный по матрице несмещенных генетических расстояний Нея (программное обеспечение TFPGA 1.3) (28). Во всех анализах учитывался доминантный характер enJSRV как генетических маркеров. Матрица межпопуляционных расстояний была обобщена в двух измерениях с использованием анализа MDS, реализованного STATISTICA ‘9.9 программный пакет (StatSoft Inc., Талса, Оклахома, США). Каждый треугольник представляет одну из 65 протестированных популяций. Обратите внимание, что на графике названы только те популяции, которые находятся за пределами основного кластера (обведены квадратом с пунктирной линией и включают подавляющее большинство пород из Африки, Азии и Европы). (B) Трехмерный график, суммирующий данные, полученные с помощью анализа главных компонентов (PCA) инсерционно-полиморфных провирусов enJSRV в 65 популяциях овец, протестированных с использованием Proc Factor статистического пакета SAS/STAT® (SAS Institute Inc, Cary NC ) по рекомендациям Кавалли-Сфорца и др. (29). Были идентифицированы четыре фактора, на долю которых приходится 86,66% вариации, с собственным значением ≥ 1. Фактор 1 (по оси X) объясняет 30,09% изменчивости и может быть интерпретирован как «фактор Северного моря», различая группу популяций, сформированных из некоторых популяций овец Великобритании и континентальной Европы (включая Данию и Тексел), и другие. Фактор 2 (по оси Y), объясняющий 23,58% вариаций, отделяющих популяцию текселей от остальных. Фактор 3 (по оси Z), объяснение 22,92% вариации и может быть интерпретирован как «фактор примитивной породы», отличающий группу популяций, образованных муфлонами и скандинавскими популяциями (включая гебридские, оркнейские и соайские популяции) от остальных. Обратите внимание, что для большей наглядности рисунка популяции, образующие основной кластер, не названы.
(29). Были идентифицированы четыре фактора, на долю которых приходится 86,66% вариации, с собственным значением ≥ 1. Фактор 1 (по оси X) объясняет 30,09% изменчивости и может быть интерпретирован как «фактор Северного моря», различая группу популяций, сформированных из некоторых популяций овец Великобритании и континентальной Европы (включая Данию и Тексел), и другие. Фактор 2 (по оси Y), объясняющий 23,58% вариаций, отделяющих популяцию текселей от остальных. Фактор 3 (по оси Z), объяснение 22,92% вариации и может быть интерпретирован как «фактор примитивной породы», отличающий группу популяций, образованных муфлонами и скандинавскими популяциями (включая гебридские, оркнейские и соайские популяции) от остальных. Обратите внимание, что для большей наглядности рисунка популяции, образующие основной кластер, не названы.
Однородные ретротипы (R2, R2-R4), которые мы наблюдали у овец современной Турции, Ирана, Саудовской Аравии, Сирии, Израиля и Египта, в сочетании с имеющимися археологическими данными позволяют предположить, что отбор домашних овец с желаемым вторичным характеристики, общие для современных пород, возникли сначала в Юго-Западной Азии, а затем успешно распространились в Европе, Африке и остальной части Азии. Это может обеспечить генетическую поддержку теории о том, что специализированное производство шерсти возникло в Юго-Западной Азии, а затем распространилось по всей Европе (11). Примитивные породы пережили вторую миграцию улучшенных пород из Юго-Западной Азии, вернувшись в дикое или полудикое состояние на острова без хищников или заняв труднодоступные районы, менее склонные к коммерческому обмену и связанной с ним интрогрессии. Большинство, если не все, породы, которые мы идентифицировали как древние, уже считались примитивными на основании таких морфологических признаков, как более темная и грубая шерсть (вместо более белого шерстяного руна), линька и частое наличие рогов. у женщин, а также мужчин ().
Это может обеспечить генетическую поддержку теории о том, что специализированное производство шерсти возникло в Юго-Западной Азии, а затем распространилось по всей Европе (11). Примитивные породы пережили вторую миграцию улучшенных пород из Юго-Западной Азии, вернувшись в дикое или полудикое состояние на острова без хищников или заняв труднодоступные районы, менее склонные к коммерческому обмену и связанной с ним интрогрессии. Большинство, если не все, породы, которые мы идентифицировали как древние, уже считались примитивными на основании таких морфологических признаков, как более темная и грубая шерсть (вместо более белого шерстяного руна), линька и частое наличие рогов. у женщин, а также мужчин ().
Открыть в отдельном окне
Морфологическая характеристика примитивных пород. Породы, идентифицированные в этом исследовании как остатки первых миграций овец, обладают морфологическими характеристиками (такими как более темная и грубая шерсть, линька, частое наличие рогов у самок), сходными с дикими овцами и муфлонами. (A) Уриальные овцы; (B) Кипрский муфлон; (C) Средиземноморский муфлон; (D) Оркнейские овцы; (E) Соевые овцы; (F) Гуте овцы; (G) Аландские овцы; (H) Исландская овца; (I) Гебридская овца.
(A) Уриальные овцы; (B) Кипрский муфлон; (C) Средиземноморский муфлон; (D) Оркнейские овцы; (E) Соевые овцы; (F) Гуте овцы; (G) Аландские овцы; (H) Исландская овца; (I) Гебридская овца.
Наше исследование также предоставляет генетические доказательства, подтверждающие анекдотическое происхождение некоторых менее распространенных пород овец. Например, одна из 10 проанализированных популяций с Британских островов, овца Джейкоб, продемонстрировала гомогенный ретротип R2, очень отличающийся от других британских популяций и более похожий на юго-западные азиатские и африканские породы. Происхождение Джейкоба неизвестно. Эта порода обязана своим названием библейской истории об Иакове, который взял «каждую крапчатую и пятнистую овцу» в качестве платы от своего тестя Лавана (Бытие 30:25-43; вероятно, первое зарегистрированное использование селекционного разведения в животноводстве). Наш ретротипический анализ подтверждает прямую связь между овцами Джейкоб и породами Юго-Западной Азии или Африки, а не другими британскими породами. Наше исследование также прочно связывает соайских овец со средиземноморскими и азиатскими муфлонами, а не с северными породами.
Наш ретротипический анализ подтверждает прямую связь между овцами Джейкоб и породами Юго-Западной Азии или Африки, а не другими британскими породами. Наше исследование также прочно связывает соайских овец со средиземноморскими и азиатскими муфлонами, а не с северными породами.
В заключение, полиморфная природа enJSRV выявила значительный вторичный рост популяции улучшенных домашних овец, скорее всего, из Юго-Западной Азии, что дает ценную информацию об истории скотоводческих обществ, экономика которых включала овцеводство. Впервые дифференцируя генетически примитивные породы от современных, наше исследование предлагает обоснование для выявления и сохранения редких генофондов. Наконец, мы демонстрируем полезность эндогенных ретровирусов как нового класса генетических маркеров, используемых для раскрытия истории одомашненных видов.
3
Нажмите здесь для просмотра. (418K, pdf)
Благодарим консорциум ECONOGENE, North-SheD и NordGen, Кэролайн Леру, Джима ДеМартини, Джеймса Траффорда, Сьюзан Хейвуд, Вальгердура Андресдоттира, Майкла Райхерта, Иоланду Байон Гонсалес, Катю Фойгт, Владимира. Feinstein, Fermin San Primitivo и Paul Halstead за помощь в получении некоторых образцов, использованных в этом исследовании, и за полезные комментарии. Мы хотели бы поблагодарить Пабло Мурсию за введение термина «ретротипы». Мы благодарны Г.П. Ди Мео и А. Перукатти за анализ FISH. Мы также благодарим Брента Хаффмана (www.ultimateungulate.com), NordGen, Свена Джеппсона и Кеннета Хедспита за фотографии, воспроизведенные в формате . Это исследование было поддержано BBSRC, Wellcome Trust, NIH Grant HD05274, Шотландским советом по финансированию через грант на развитие стратегических исследований и частично от Fundação para a Ciência e a Tecnologia, «Misura P5 Biodiversita’ animale» от Regione autonoma della Sardegna. и Китайской академии наук (KSCX2-YW-N-018). Проект по разведению сойских овец в Сент-Килде поддерживается NERC. IPATIMUP частично поддерживается «Programa Operacional Ciência e Inovação 2010» (POCI 2010), VI Programa Quadro (2002–2006). Член парламента является лауреатом премии Wolfson-Royal Society Research Merit.
Feinstein, Fermin San Primitivo и Paul Halstead за помощь в получении некоторых образцов, использованных в этом исследовании, и за полезные комментарии. Мы хотели бы поблагодарить Пабло Мурсию за введение термина «ретротипы». Мы благодарны Г.П. Ди Мео и А. Перукатти за анализ FISH. Мы также благодарим Брента Хаффмана (www.ultimateungulate.com), NordGen, Свена Джеппсона и Кеннета Хедспита за фотографии, воспроизведенные в формате . Это исследование было поддержано BBSRC, Wellcome Trust, NIH Grant HD05274, Шотландским советом по финансированию через грант на развитие стратегических исследований и частично от Fundação para a Ciência e a Tecnologia, «Misura P5 Biodiversita’ animale» от Regione autonoma della Sardegna. и Китайской академии наук (KSCX2-YW-N-018). Проект по разведению сойских овец в Сент-Килде поддерживается NERC. IPATIMUP частично поддерживается «Programa Operacional Ciência e Inovação 2010» (POCI 2010), VI Programa Quadro (2002–2006). Член парламента является лауреатом премии Wolfson-Royal Society Research Merit.
Вспомогательный онлайн-материал
www.sciencemag.org
Материалы и методы
Рис. S1, S2, S3
Таблицы S1, S2
1. Зедер М.А. Proc Natl Acad Sci U S A. 2008; 105:11597. [Бесплатная статья PMC] [PubMed] [Google Scholar]
2. College S, Conolly J, Shennan S. European Journal of Archaeology. 2005; 8:137. [Google Scholar]
3. Цена ТД. Первые фермеры Европы. Издательство Кембриджского университета; Кембридж: 2000. [Google Scholar] 9.0011
4. Баркер Г. В кн.: Исследование гипотезы распространения сельского хозяйства/языка. Беллвуд П., Ренфрю С., редакторы. Монографии Института Макдональда; Кембридж: 2002. стр. 151–162. [Google Scholar]
5. Харрис Д.Р., Госден К. В кн.: Происхождение и распространение сельского хозяйства и скотоводства в Евразии. Харрис Д., редактор. УКЛ пресс; Лондон: 1996. С. 370–389. [Google Scholar]
6. Райдер М.Л. Овца и человек. Джеральд Дакворт и Ко; London: 1983. [Google Scholar]
7. Pedrosa S, et al. Proc Biol Sci. 2005; 272:2211. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Pedrosa S, et al. Proc Biol Sci. 2005; 272:2211. [Бесплатная статья PMC] [PubMed] [Google Scholar]
8. Tapio M, et al. Мол Биол Эвол. 2006; 23:1776. [PubMed] [Google Scholar]
9. Медоуз Дж. Р., Джемаль И., Караджа О., Гутвайн Э., Киджас Дж. В. Генетика. 2007; 175:1371. [Бесплатная статья PMC] [PubMed] [Google Scholar]
10. Naderi S, et al. Proc Natl Acad Sci U S A. 2008; 105:17659. [Бесплатная статья PMC] [PubMed] [Google Scholar]
11. Шерратт А. В: Образцы прошлого: Исследования в честь Дэвида Кларка. Ходдер И., Исаакс Г., Хаммонд Н., редакторы. Издательство Кембриджского университета; Кембридж: 1981. [Google Scholar]
12. Helmer D, Gourichon L, Vila E. Anthropozoologica. 2007;42:41. [Google Scholar]
13. Бёке Дж.Д., Стойе Дж.П. В: Ретровирусы. Гроб Дж. М., Хьюз С. Х., Вармус Х. Е., редакторы. Лабораторный пресс Колд-Спринг-Харбор; Плейнвью, Нью-Йорк: 1997. стр. 343–436. [Google Scholar]
14. Arnaud F, et al. Возбудители PLoS. 2007;3:e170. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
15. Palmarini M, Sharp JM, De las Heras M, Fan H. Journal of Virology. 1999;73:6964. [Бесплатная статья PMC] [PubMed] [Google Scholar]
16. Arnaud F, Varela M, Spencer TE, Palmarini M. Cell Mol Life Sci. 2008 [бесплатная статья PMC] [PubMed] [Google Scholar]
17. Материалы и методы доступны в качестве вспомогательного материала на Science Online.
18. Эрнандес Фернандес М., Врба Э.С. Biol Rev Camb Philos Soc. 2005; 80:269. [PubMed] [Google Scholar]
19. Hiendleder S, Kaupe B, Wassmuth R, Janke A. Proc Biol Sci. 2002; 269:893. [Бесплатная статья PMC] [PubMed] [Google Scholar]
20. Клаттон-Брок Дж. Естественная история домашних млекопитающих. изд. 2-е издательство Кембриджского университета; Cambridge: 1999. [Google Scholar]
21. Зохари Д., Чернов Э., Хорвиц Л. Зоологический журнал. 1998; 245:129. [Google Scholar]
22. Поплин Ф. Анналы генетики и селекции животных. 1979; 11:133. [Бесплатная статья PMC] [PubMed] [Google Scholar]
23. Vigne J-D. В: Голоценовая история европейской позвоночной фауны. Современные аспекты исследования. Бенеке Н., редактор. Verlag Marie Leidorf GmbH, Раден/Вестф; Германия: 1999. стр. 295–322. [Google Scholar]
24. Pedrosa S, et al. Генет Сель Эвол. 2007; 39:91. [Бесплатная статья PMC] [PubMed] [Google Scholar]
25. Pereira F, et al. Мол Биол Эвол. 2006; 23:1420. [PubMed] [Google Scholar]
26. McGovern TH. Ежегодный обзор антропологии. 1990;19:331. [Google Scholar]
27. Экскофье Л., Лаваль Г., Шнайдер С. Эволюционная биоинформатика в Интернете. 2005; 1:47. [Бесплатная статья PMC] [PubMed] [Google Scholar]
28. Miller MP. Департамент биологических наук Университета Северной Аризоны; Флагстафф, Аризона: 1997. [Google Scholar]
29. Кавалли-Сфорца Л.Л., Меноцци П., Пьяцца А. История и география генов человека. Издательство Принстонского университета; Принстон, США: 1994. [Google Scholar]
[Google Scholar]
| Раннее одомашнивание Овцы в Новом Свете В 1519 году Кортес начал свое исследование Мексики и Западного
Соединенные Штаты. Побережье Мексиканского залива (или Флорида) Родной — еще одна порода овец. считается прямым потомком овец, завезенных в Новый Мир испанскими и французскими исследователями. Ферал до начала 20-х века, аборигенные овцы побережья Мексиканского залива известны своей естественной устойчивостью к глистным паразитам. Ранняя американская история К 1698 году Америка экспортировала изделия из шерсти. Англия была возмущена
и объявили торговлю шерстью вне закона, сделав ее наказуемой отрезанием шерсти.
правая рука человека. Ограничения на разведение овец и шерсти
производство, наряду с Законом о гербовых марках, привело к американскому революционному
Война. Таким образом, прядение и ткачество считались патриотическими поступками.
Даже после войны в Англии был принят закон, запрещающий экспорт
из любых овец, но wethers. В конфликте Во время пастбищных войн в США вспыхнули ожесточенные конфликты между скотом
владельцы ранчо и пастухи, поскольку оба конкурировали за землю для выпаса скота
их домашний скот. Закрытие общего пользования в Великобритании было похоже на
клиренс Хайленд; открытые поля были заключены в индивидуальные владения
поля для овцеводства, вытеснив многих фермеров, ведущих натуральное хозяйство. |
 до н.э. что люди научились прясть шерсть.
Овцы помогли сделать возможным распространение цивилизации. Овца
производство было хорошо налажено в библейские времена. Есть
много ссылок на овец в Библии, особенно в Ветхом Завете.
Овцеводство — старейшая организованная отрасль человека. Шерсть была
первый товар достаточной стоимости, чтобы гарантировать международное
торговля.
до н.э. что люди научились прясть шерсть.
Овцы помогли сделать возможным распространение цивилизации. Овца
производство было хорошо налажено в библейские времена. Есть
много ссылок на овец в Библии, особенно в Ветхом Завете.
Овцеводство — старейшая организованная отрасль человека. Шерсть была
первый товар достаточной стоимости, чтобы гарантировать международное
торговля. Он взял с собой овец, потомков Колумба.
овца. Считается, что эти овцы являются потомками того, что
теперь называются «Чуррос». Навахо чурро — старейший
порода овец в США. Несмотря на усилия правительства США
чтобы искоренить породу, навахо по-прежнему выращивают чуррос навахо.
индейцы.
Он взял с собой овец, потомков Колумба.
овца. Считается, что эти овцы являются потомками того, что
теперь называются «Чуррос». Навахо чурро — старейший
порода овец в США. Несмотря на усилия правительства США
чтобы искоренить породу, навахо по-прежнему выращивают чуррос навахо.
индейцы. Инаугурационная куртка Джеймса Мэдисона была соткана из овечьей шерсти.
вырос в своем доме в Вирджинии. Президент Вудро Вильсон пасся
овцы на лужайке Белого дома.
Инаугурационная куртка Джеймса Мэдисона была соткана из овечьей шерсти.
вырос в своем доме в Вирджинии. Президент Вудро Вильсон пасся
овцы на лужайке Белого дома.