Гидрохимическая характеристика реки Обь
- Подробности
- Категория: Великая сибирская река Обь
- Просмотров: 1151
Согласно классификации О.А. Алекинаводы р. Обь относятся к гидрокарбонатному классу вод группы кальция. Показатели величин рН лежат в области слабощелочных значений, величина окислительно-восстановительного потенциала принимает положительные значения, высокий уровень содержания растворенного кислорода. Воды р. Обь обладают средней жесткостью и минерализацией.
Биогенные элементы в водах р. Обь находятся на допустимом уровне, но в период весеннего половодья отмечаются превышения допустимых значений для ионов Nh5+ и РО4+3, что можно объяснить поступлением этих ионов с водосборной площади во время снеготаяния. Воды Оби характеризуются большим содержанием органических веществ и низким содержанием кислорода, что зимой приводит к заморам. Средняя мутность снижается вниз по течению от 160 до 40 г/м [2].
Для территории Обь-Иртышского бассейна выделяют 5 зон по характеру минерализации и химического состава, представленным показателем комбинации трех ионов, содержание которых преобладает в солевом составе воды [5]:
1. Тундрово-лесотундровая – зона преобладания гидрокарбонатно-кремнеземных, богатых органическим веществом гидрохимических фаций (НСО3-SiО2-SО4, SiО2- НСО3-Са, НСО3-Са- SiО2 и др.). Здесь водные массы имеют малую минерализацию и жесткость, большую окисляемость и цветность, которая придает воде желто-коричневую окраску из-за высокого содержания растворенных в воде органических веществ и оксидов железа, выносимых с мерзлых грунтов и заболоченных водосборов.
2. Таежная – зона преобладания гидрокарбонатно-кальциевых гидрохимических фаций (НСО3-Са-SО4, НСО3-Са-Мg, НСО3-Са-Na и др.). Средняя минерализация здесь примерно вдвое выше, а цветность имеет повышенную величину лишь в половодье.

3. Степная – зона преобладания гидрокарбонатно-сульфатных, сульфатных и хлоридных гидрохимических фаций (НСО3-SО4-Са, SО4-НСО3-Сl, Cl-SО4-НСО3 и др.). Водные массы имеют среднюю минерализацию, вдвое большую по сравнению с водами лесной зоны.
4. Области внутреннего стока – зона преобладания хлоридно-натриевых гидрохимических фаций (Cl-Nа-SО4, Cl-SО4-НСО3, SО4-Сl–Nа и др.). Повышенная минерализация, трансформация химического состава воды.
5. Горные области – зона преобладания вертикальных подзон: преобладания кремнеземных (SiО2-НСО3-Са) и гидрокарбонатно-кремнеземных (НСО3-SiО2-Са) фаций альпийских лугов, преобладания гидрокарбонатных (НСО3-Са-SО4, НСО3-SО4-Са, СО3-SО4-Nа) фаций горных лесов и преобладания сульфатных фаций предгорных степей (SО4-НСО3-Nа). Полизональность горных рек проявляется не столько в изменении водного режима с высотой, сколько в гидрохимической трансформации их водных масс при смешении с водами притоков, формирующими сток в различных высотных поясах.
Из природных источников наибольшее влияние на химический состав обского бассейна оказывает почвенный покров водосборов.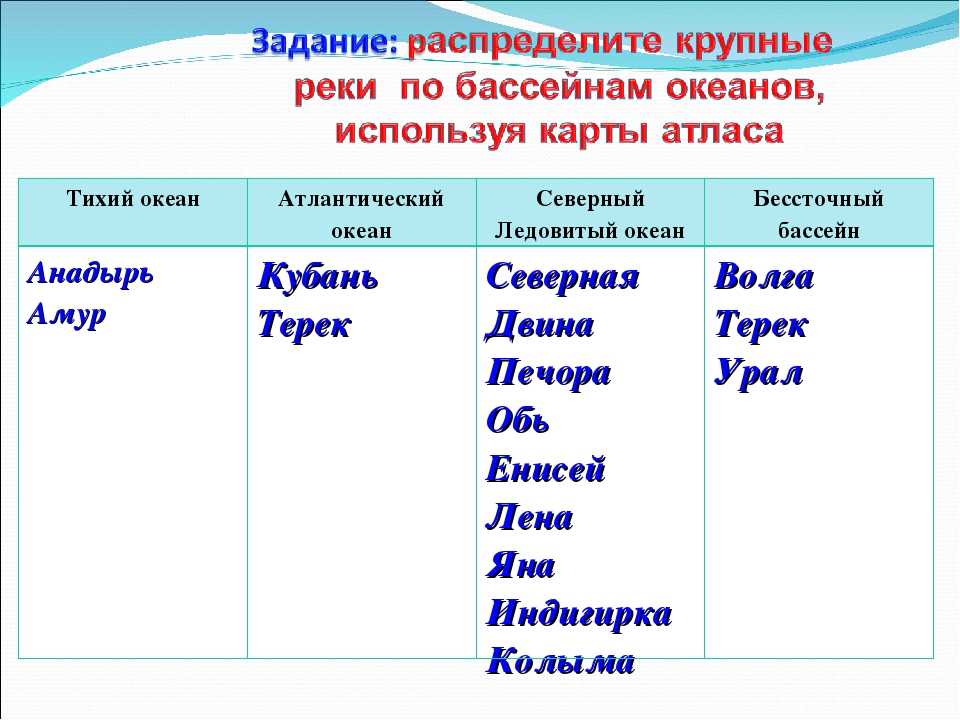 Геохимическая обстановка меняется в пределах от окислительной слабощелочной и щелочной в повышенных аридных элементах рельефа до восстановительной глеевой слабокислой – в пониженных, избыточно увлажненных и на участках развития слабопроницаемых пород (глин, суглинков). В формировании гидрохимического состава вод Оби особую роль играет наличие большого количества органического вещества в опаде растительного покрова водосборов и почвах. Почвы водохозяйственных участков определяют ионный состав поверхностных вод в летний период, в связи с повышенным проявлением биогеохимических процессов и выраженным поверхностным и внутрипочвенным геохимическим стоком [5].
Геохимическая обстановка меняется в пределах от окислительной слабощелочной и щелочной в повышенных аридных элементах рельефа до восстановительной глеевой слабокислой – в пониженных, избыточно увлажненных и на участках развития слабопроницаемых пород (глин, суглинков). В формировании гидрохимического состава вод Оби особую роль играет наличие большого количества органического вещества в опаде растительного покрова водосборов и почвах. Почвы водохозяйственных участков определяют ионный состав поверхностных вод в летний период, в связи с повышенным проявлением биогеохимических процессов и выраженным поверхностным и внутрипочвенным геохимическим стоком [5].
- Назад
- Вперёд
Власиха (река) — справка
Главная » Реки
Реки
Власиха — малая река в Алтайском крае, левый приток Барнаулки. Бассейн реки частично находится в пределах города Барнаула в его юго-западной части.
Бассейн реки частично находится в пределах города Барнаула в его юго-западной части.
| Справочные данные | |
|---|---|
| Длина | 18 км |
| Бассейн | 119 км² |
| Расход воды | 1 м³/с |
| Исток | |
| Высота | 240 м |
| Устье | Барнаулка |
| 18 км от устья по левому берегу | |
| Координаты | 53°16′30″ с. ш. 83°37′12″ в. д. |
| Водная система | Барнаулка → Обь → Карское море |
| Страна |
|
| Регион | Алтайский край |
| Район | Барнаул |
| Код в ГВР | 13010200512115200001335 |
Река Власиха протекает по Приобскому плато с севера-запада на юго-восток. Исток находится у посёлка Шахи Павловского района Алтайского края на высоте около 240 м над уровнем моря, но из-за нерегулярности стока часть русла реки пересыхает. Регулярный сток образуется после запруженной части реки — севернее одноимённого реке села Власиха. Впадает Власиха в Барнаулку, на 2 км выше по течению от посёлка Борзовая Заимка на высоте 150 м.
Регулярный сток образуется после запруженной части реки — севернее одноимённого реке села Власиха. Впадает Власиха в Барнаулку, на 2 км выше по течению от посёлка Борзовая Заимка на высоте 150 м.
Модуль годового стока колеблется от 1 до 5 л/с с км², а расход воды составляет — менее 1 м³/с. Общий объём годового стока около 3,5 млн м³. Тип водного режима относится рекам с весенним половодьем и редкими паводками в летне-осенний период. Снеговое и дождевое питание — 80-85 %, грунтовое — 15—20 %. Площадь водосборного бассейна — 119 км².
В пределах барнаульского ленточного бора река пересекает территорию активного оврагообразования. Окружающие ландшафты представлены днищами балок с лугово-кустарниковой растительностью на аллювиально-луговых почвах, а также придолинными склонами с остепнённой луговой растительностью. При этом данные ландшафты имеют средний эколого-природный потенциал и высокую антропогенную нагрузку.
Инженерно-геологические условия представлены современными отложениями пролюво-делювия: песками, суглинками и супесями.
Река Власиха используется местным населением прежде всего как источник пресной питьевой воды, так как долина реки является водоносным комплексом. В верхнем течении это воды спорадического распространения в нижне-среднечетвертичных отложениях краснодубровской свиты, а в нижнем — водоносные горизонты верхнечетвертичных аллювиальных отложений надпойменных террас Барнаулки.
Выше по течению от села Власиха, долина реки активно используется в качестве пастбища и сенокоса, а основная часть реки, пересекающая ленточный бор в рекреационных целях: здесь находятся летние детские оздоровительные лагеря, места для загородного отдыха горожан.
По данным государственного водного реестра России относится к Верхнеобскому бассейновому округу, водохозяйственный участок реки — Обь от города Барнаул до Новосибирского гидроузла, без реки Чумыш, речной подбассейн реки — бассейны притоков (Верхней) Оби до впадения Томи. Речной бассейн реки — (Верхняя) Обь до впадения Иртыша.
Источник
Россия Реки России Реки Алтайского края Реки Алтайский крайВодный режим почв и водный баланс
Водный режим почвы и водный баланс. Совокупность всех процессов поступления воды в почву, состояния воды в почве и водопотребления в почве называется водным режимом. Количественное определение всех видов притока и отвода воды в почву за определенный интервал времени называется водным балансом.
Совокупность всех процессов поступления воды в почву, состояния воды в почве и водопотребления в почве называется водным режимом. Количественное определение всех видов притока и отвода воды в почву за определенный интервал времени называется водным балансом.
Водный режим почвы — это совокупность всех явлений поступления воды в почву, ее движения, изменения ее физического состояния и отвода из почвы.
Основы теории водного режима почв были заложены Г.Н. Высоцкий (1934) и А.А. Роде (1956). Они разбили водный режим почвы на шесть типов и несколько подтипов. Позднее работы И.А. Качинский (1970), И.Г. Минашина (1974), М.А. Козин (1977), А.Г. Бондарев (1996), О.И. Худяков (1988) и др. были посвящены решению этой проблемы.
К компонентам водного режима относятся абсорбция, инфильтрация, капиллярный подъем, поверхностные, нисходящие и боковые течения, физическое испарение, замерзание, размораживание и конденсация воды.
В зависимости от количественного соотношения между этими явлениями различают следующие типы водного режима (по Роде):
- Криогенный режим характерен для районов вечной мерзлоты.
 Всегда имеется водоупорная толща (аквифуга). В безморозные периоды почва насыщается влагой за счет образования торчащих вод. Отношение осадков к испаряемости PER > 1. Характерно для арктических и тундровых почв.
Всегда имеется водоупорная толща (аквифуга). В безморозные периоды почва насыщается влагой за счет образования торчащих вод. Отношение осадков к испаряемости PER > 1. Характерно для арктических и тундровых почв. - Выщелачивающий режим характерен для районов с отношением осадков к испаряемости PER > 1, т.е. сумма осадков больше испаряемости. Атмосферная влага проникает через всю почвенную толщу и достигает грунтовых вод. Вымываются щелочные и щелочноземельные элементы (подзолы/сподосоли, бурые лесные почвы, красноземы/красноземы, желтоземы/желтоземы). Подтип заболоченных почв развивается при PER > 1, неглубоких грунтовых водах и наличии водоупоров (подзолистые болотные и заболоченные почвы).
- Периодически-промывной режим: PER = 1. Характеризуется чередованием непромывного и промывного режимов воды в засушливые и многоводные годы (серые лесные почвы, оподзоленные почвы и выщелоченные черноземы/черноземы).
- Безпромывочный режим: PER < 1. Атмосферная влага распространяется в верхние горизонты почвы и никогда не достигает грунтовых вод. В степной почве влага проникает на глубину 3-4 м; на пустынных почвах (бурых пустынно-степных и серо-бурых пустынных) до 1 м.
- Экссудационный режим: PER < 1. Характерен для степных, полупустынных и пустынных территорий с неглубоким залеганием грунтовых вод. Здесь преобладают вертикальные течения. При высокой минерализации грунтовых вод происходит засоление почв, развиваются солонцеватые почвы. При десуктивно-экссудационном режиме растения перехватывают воду, и влага испаряется за счет транспирации, соли откладываются в толще, а не на поверхности.
- Режим орошения создается при проведении орошения. Для него характерно чередование промывных и непромывных или даже экссудационных режимов воды.
 Всегда имеется водоупорная толща (аквифуга). В безморозные периоды почва насыщается влагой за счет образования торчащих вод. Отношение осадков к испаряемости PER > 1. Характерно для арктических и тундровых почв.
Всегда имеется водоупорная толща (аквифуга). В безморозные периоды почва насыщается влагой за счет образования торчащих вод. Отношение осадков к испаряемости PER > 1. Характерно для арктических и тундровых почв. В степной почве влага проникает на глубину 3-4 м; на пустынных почвах (бурых пустынно-степных и серо-бурых пустынных) до 1 м.
В степной почве влага проникает на глубину 3-4 м; на пустынных почвах (бурых пустынно-степных и серо-бурых пустынных) до 1 м.Водный режим выражается через водный баланс, характеризующийся поступлением и расходом воды. По Роде, водный баланс почвы за определенный период выражается в общем виде следующим уравнением:
M 1 = M 0 + (R + K + GW) — (T + E + FS + FL + FG)
Где:
М 1 – запас влаги в почвенной толще на конец исследуемого периода.
М 0 – запас влаги в почвенной толще на начало изучаемого периода.
R означает общее количество осадков.
обозначает конденсацию влаги
ВВ – содержание влаги, поступающей в почву из подземных вод.
означает транспирацию
означает испарение
FS означает поверхностный поток
FL означает подземный боковой сток.
FG – сток подземных вод (в мм или м 3 /га).
Осадки и подземные воды считаются основным источником водного баланса. В то время как дополнительными источниками увлажнения почвы являются поверхностный приток и влага, конденсируемая из водяного пара. Водоотдача складывается из физического испарения воды с поверхности воды, потерь влаги при транспирации, потерь воды при поверхностных и подповерхностных боковых притоках и инфильтрации воды в толщу почвы.
Контроль водного режима почвы
Основными методами регулирования водного режима являются осушение земель, орошение, чистый пар, а также приемы, направленные на снижение непродуктивного почвенного испарения и снегозадержания.
В засушливых районах наиболее эффективным способом улучшения водного режима почвы является орошение.
Количество поливных работ, поливные и поливные нормы следует корректировать в зависимости от текущих и ожидаемых погодных условий, чтобы максимально эффективно использовать оросительную воду, запасы которой ограничены.
В субгумидной зоне чистый пар способствует накоплению влаги. К осеннему севу по чистым парам в сухостепных районах продуктивной влаги накапливается на 50-60 мм больше, чем на незалежных полях. Часто эта разница доходит даже до 70-100 мм. Большое значение для регулирования влажности почвы имеет ранняя вспашка зяблевой пахоты.
Все мероприятия, направленные на снижение непродуктивного испарения и снегозадержания в засушливой зоне степи, способствуют оптимизации водного режима почвы.
Прогнозирование воздействия изменений водного режима на водоплавающих птиц: анализ постановки лебедей
1. Pimm SL, Jenkins CN, Abell R, Brooks TM, Gittleman JL, Joppa LN, et al. Биоразнообразие видов и темпы их исчезновения, распространения и защиты. Наука. 2014;344(6187):1246752
10.1126/наука.1246752 . [PubMed] [CrossRef] [Google Scholar]
Биоразнообразие видов и темпы их исчезновения, распространения и защиты. Наука. 2014;344(6187):1246752
10.1126/наука.1246752 . [PubMed] [CrossRef] [Google Scholar]
2. Ellis EC. Экология в антропогенной биосфере. Экологические монографии. 2015;85(3):287–331. 10.1890/14-2274.1 [CrossRef] [Академия Google]
3. Гимеси А., Ван Роой Е.П., Нолет Б.А. Нелинейные эффекты агрегации пищи на интерференционную конкуренцию у крякв. Behav Ecol Sociobiol. 2010;64(11):1897–904. [Бесплатная статья PMC] [PubMed] [Google Scholar]
4. Mori Y, Boyd IL. Поведенческая основа нелинейных функциональных реакций и оптимального поиска пищи у южных морских котиков. Экология. 2004;85(2):398–410.: ISI:000220108700011. [Google Scholar]
5. Leirs H, Stenseth NC, Nichols JD, Hines JE, Verhagen R, Verheyen W. Стохастическая сезонность и нелинейные факторы, зависящие от плотности, регулируют размер популяции африканских грызунов. Природа. 1997;389:176–80. [PubMed] [Google Scholar]
6. Стиллман Р.А., Goss-Custard JD, West AD, Durell S, McGrorty S, Caldow RWG и соавт.
Прогнозирование смертности куликов и размера популяции при различных режимах управления промыслом моллюсков. J Appl Ecol. 2001;38(4):857–68.: ISI:000170568000015. [Google Scholar]
Стиллман Р.А., Goss-Custard JD, West AD, Durell S, McGrorty S, Caldow RWG и соавт.
Прогнозирование смертности куликов и размера популяции при различных режимах управления промыслом моллюсков. J Appl Ecol. 2001;38(4):857–68.: ISI:000170568000015. [Google Scholar]
7. Стиллман Р.А., Goss-Custard JD. Индивидуальная экология прибрежных птиц. Биол Rev. 2010; 85: 413–34. 10.1111/j.1469-185X.2009.00106.x [PubMed] [CrossRef] [Академия Google]
8. Стиллман Р.А., Рейлсбак С.Ф., Гиске Дж., Бергер У., Гримм В. Делать прогнозы в меняющемся мире: преимущества индивидуальной экологии. Бионаука. 2014;65(2):140–50. 10.1093/biosci/biu192 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
9. Каранек А.Р., Ламберсон Р.Х., Блэк Дж.М. Индивидуальная модель традиционного поведения при поиске пищи: исследование последствий колебаний окружающей среды. Моделирование природных ресурсов. 2008;21(1):93–116. [Google Scholar]
10. Стиллман Р.А., Вуд К.А., Гилкерсон В., Элкинтон Э. , Блэк Дж.М., Уорд Д.Х. и другие.
Прогнозирование последствий изменения окружающей среды для мигрирующих травоядных. Экосфера. 2015;6(7):арт114
10.1890/es14-00455.1 [CrossRef] [Google Scholar]
, Блэк Дж.М., Уорд Д.Х. и другие.
Прогнозирование последствий изменения окружающей среды для мигрирующих травоядных. Экосфера. 2015;6(7):арт114
10.1890/es14-00455.1 [CrossRef] [Google Scholar]
11. Van Eerden MR, De Leeuw JJ, Slager B, Bij de Vaate A. Полевые испытания концепции грузоподъемности зимующих ныряющих уток: ограничивают ли высокие затраты на кормление эксплуатация мидий-зебр? В: Van Eerden MR, редактор. Пэчворк. Лелистад: Directie IJsselmeergebied, Rijkswaterstaat; 1997. с. 283–316. [Google Scholar]
12. Goss-Custard JD, Stillman RA, West AD, Caldow RWG, Triplet P, Durell S, et al. Когда недостаточно: кулики и ловля моллюсков. Труды Королевского общества, Лондон Б. 2004; 271 (1536): 233–7 .: ISI: 00018869.4400003. [Бесплатная статья PMC] [PubMed] [Google Scholar]
13. Williams CK, Dugger BD, Brasher MG, Collucy JM, Cramer DM, Eadie JM, et al.
Оценка пропускной способности среды обитания для мигрирующих и зимующих водоплавающих птиц: соображения, подводные камни и улучшения. дичь. 2014;4
Специальный выпуск: 407–35. [Google Scholar]
дичь. 2014;4
Специальный выпуск: 407–35. [Google Scholar]
14. Ловворн Дж.Р. Биомеханика и рентабельность кормодобывания: подход к оценке трофических потребностей и воздействия ныряющих уток. гидробиол. 1994; 279/280: 223–33. [Академия Google]
15. Де Леу Дж. Дж. Затраты на ныряние как компонент дневного энергетического баланса водных птиц и млекопитающих: обобщение включения затрат на восстановление после погружения, продемонстрированное у хохлатых уток. Джан Джей Зул. 1996; 74: 2131–42. [Google Scholar]
16. Гавлик Д.Э. Влияние доступности добычи на числовую реакцию болотных птиц. Эколь моногр. 2002; 72: 329–46. [Google Scholar]
17. Gyimesi A, Varghese S, de Leeuw J, Nolet BA. Коэффициент чистого потребления энергии как общепринятая валюта для объяснения пространственного распределения лебедей в мелководном озере. водно-болотные угодья. 2012;32:119–27. 10.1007/s13157-011-0256-6 [CrossRef] [Google Scholar]
18. Clausen P. Моделирование влияния уровня воды на выбор среды обитания и доступность пищи для Zostera , кормящих черных казарок Branta bernicla в неприливных районах. Биология дикой природы. 2000; 6: 75–87. [Google Scholar]
Биология дикой природы. 2000; 6: 75–87. [Google Scholar]
19. Алатало Р.В., Эрикссон Д., Густафссон Л., Ларссон К. Конкуренция при эксплуатации влияет на использование синичками мест кормления — экспериментальные данные. Экология. 1987;68(2):284–90.: ISI:A1987G467000006. [Академия Google]
20. Браун Дж.С. Использование участка как показатель предпочтения среды обитания, риска хищничества и конкуренции. Behav Ecol Sociobiol. 1988; 22:37–47. [Google Scholar]
21. Anholt BR. Экспериментальное разделение вмешательства и эксплуататорской конкуренции у личиночной стрекозы. Экология. 1990;71(4):1483–93.: ISI:A1990DQ19100028. [Google Scholar]
22. Caldow RWG, Goss-Custard JD, Stillman RA, Le V, Dit Durell SEA, Swinfen R, et al. Индивидуальные различия в конкурентоспособности склонных к помехам собирателей: относительная важность эффективности поиска пищи и восприимчивости к помехам. Дж Аним Экол. 1999;68:869–78. [Google Scholar]
23. Grimm V, Revilla E, Berger U, Jeltsch F, Mooij WM, Railsback SF, et al. Шаблонно-ориентированное моделирование сложных агентных систем: уроки экологии. Наука. 2005; 310(5750):987–91.: ISI:000233343400034. [PubMed] [Google Scholar]
Шаблонно-ориентированное моделирование сложных агентных систем: уроки экологии. Наука. 2005; 310(5750):987–91.: ISI:000233343400034. [PubMed] [Google Scholar]
24. Logemann D, Klasberg M, Kampkuiper A, Van Leussen J. Beheeren inrichtringsplan National Park Lauwersmeer. Assen: Arcadis, 2003. [Google Scholar]
25. Beekman JH, Van Eerden MR, Dirksen S, Bewick’s Swans. Cygnus columbianus bewickii использует меняющийся ресурс Potamogeton pectinatus осенью в Нидерландах. дичь. 1991; 1 Приложение: 238–48. [Google Scholar]
26. Нолет Б.А., Андреев В.А., Клаузен П., Пут М.Дж.М., Вессель Э.Дж. Значение Белого моря как остановки для весенних лебедей Cygnus columbianus bewickii . Ибис. 2001;143(1):63–71.: ISI:000166159100008. [Google Scholar]
27. Нолет Б.А., Фулд В.Н., Van Rijswijk MEC. Затраты на добычу корма и доступность как факторы, определяющие отказ от плотности в системе с рдестами. Ойкос. 2006; 112: 353–62. [Google Scholar]
28. Nolet BA, Langevoord O, Bevan RM, Engelaar KR, Klaassen M, Mulder RJW, et al. Пространственные различия в истощении клубней лебедями объясняются различиями в чистой скорости потребления. Экология. 2001; 82 (6): 1655–67 .: ISI: 000169.521200013. [Google Scholar]
Пространственные различия в истощении клубней лебедями объясняются различиями в чистой скорости потребления. Экология. 2001; 82 (6): 1655–67 .: ISI: 000169.521200013. [Google Scholar]
29. Нолет Б.А., Дрент Р.Х. Малые лебеди заправляются клубнями рдеста в Двинском заливе (Белое море) во время весенней миграции: в порядке живой очереди. Дж. Авиан Биол. 1998;29(4):574–81.: ISI:000078186100025. [Google Scholar]
30. Гимеси А., Стиллман Р.А., Нолет Б.А. Соревнование по скрытой интерференции у лебедей, добывающих загадочную добычу. Поведение животных. 2010;80(5):791–7. [Google Scholar]
31. Нолет Б.А., Гимеси А., Клаассен Р.Х.Г. Прогнозирование пропускной способности птичьего дня на промежуточной площадке: проверка моделей истощения. Дж Аним Экол. 2006; 75: 1285–9.2. [PubMed] [Google Scholar]
32. Нолет Б.А., Клаассен М. Ретродиктирование использования пластыря при поиске пищи лебедями в гетерогенной среде с использованием набора функциональных ответов. Ойкос. 2009; 118: 431–9. [Google Scholar]
2009; 118: 431–9. [Google Scholar]
33. Нолет Б.А., Гимеси А. Недостаточное использование места остановки мигрирующими лебедями. Дж Орнитол. 2013; 154: 695–703. 10.1007/s10336-013-0934-7 [CrossRef] [Google Scholar]
34. Нолет Б.А., Беван Р.М., Клаассен М., Лангеворд О., Хейден В.Д. Переключение среды обитания лебедями Бьюика: максимизация среднего долгосрочного прироста энергии. Дж Аним Экол. 2002;71:979–93. [Google Scholar]
35. Нолет Б.А., Клаассен М. Ограничение времени и энергии в трудные фазы годового цикла: пример ограничения времени при дозаправке перелетных лебедей. Ойкос. 2005; 111: 302–10. [Google Scholar]
36. Клаассен Р.Х.Г., Нолет Б.А., Банкерт Д. Движение тундровых лебедей, собирающих корм, объясняется пространственной структурой загадочной плотности пищи. Экология 2006;87(9):2244–54. [PubMed] [Google Scholar]
37. Норберг У. Энергетика полета В: Carey C, редактор. Энергетика птиц и экология питания. Нью-Йорк: Чепмен и Холл; 1996. с. 199–249. [Google Scholar]
199–249. [Google Scholar]
38. Бикман Дж. Х., Бертольд П., Новак Э., Квернер Ю. Внедрение спутникового слежения при изучении миграции утиных: обзор и тематическое исследование. Гибье Фаун Соваж. 1996; 13: 157–76. [Google Scholar]
39. Мэдсен Дж., Клаассен М. Оценка состояния тела и компонентов энергетического баланса путем подсчета профилей брюшной полости у живущих на свободе розовых гусей Anser brachyrhynchus. Дж. Авиан Биол. 2006; 37: 283–7. [Google Scholar]
40. Боулер Дж.М. Состояние малых лебедей Cygnus columbianus bewickii зимой, оцененное по профилям их брюшка. Ардея. 1994;82:241–8. [Google Scholar]
41. Ван Эрден М.Р., Бикман Дж.Х., Смит М., Остербик К. Использование участков лебедями Бьюика Cygnus columbianus bewickii, питающимися саговым рдестом Potamogeton pectinatus в мелководных озерах Нидерландов: различия в пороговых значениях эксплуатации, вызванные социальными, Факторы, зависящие от окружающей среды и времени. В: Ван Эрден М.Р., редактор. Пэчворк. Lelystad: Rijkswaterstaat, Directie IJsselmeergebied; 1997. с. 187–214. [Google Scholar]
Пэчворк. Lelystad: Rijkswaterstaat, Directie IJsselmeergebied; 1997. с. 187–214. [Google Scholar]
42. Пот Р. Развитие потамогетонной растительности в Лауверсмере. Акта Бот Нирл. 1984;33:361–2. [Google Scholar]
43. Hidding B, Nolet BA, de Boer T, de Vries PP, Klaassen M. Наземные и подземные травоядные позвоночные могут отдавать предпочтение различным подчиненным видам в сообществе водных растений. Экология. 2010; 162:199–208. 10.1007/s00442-009-1450-6 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
44. Van Wijk RJ. Экологические исследования Potamogeton pectinatus Л.И. Общие характеристики, производство биомассы и жизненные циклы в полевых условиях. Аква Бот. 1988;31:211–58. [Google Scholar]
45. Brouwer GA, Tinbergen L. De verspreiding der Kleine Zwanen, Cygnus b. bewickii Yarr., in de Zuiderzee, vóór en na de verzoeting. Лимоза. 1939; 12: 1–18 (перепечатано в Limosa 75: 2–12). [Google Scholar]
46. Гимеси А. , Ван Лит Б., Нолет Б.А. Комменсальное кормление лебедями Бьюика Cygnus bewickii удваивает мгновенную скорость потребления обыкновенных нырков Aythya ferina. Ардея. 2012; 100:55–62. [Google Scholar]
, Ван Лит Б., Нолет Б.А. Комменсальное кормление лебедями Бьюика Cygnus bewickii удваивает мгновенную скорость потребления обыкновенных нырков Aythya ferina. Ардея. 2012; 100:55–62. [Google Scholar]
47. Нолет Б.А., Беван Р.М., Клаассен М., Лангеворд О., Ван дер Хейден YGJT. Смена среды обитания лебедями Бьюика: максимизация среднего долгосрочного прироста энергии? Дж Аним Экол. 2002;71:979–93. [Google Scholar]
48. Нолет Б.А., Муий В.М. Поисковые пути лебедей, кормящихся на пространственно автокоррелированных клубнях. Дж Аним Экол. 2002; 71: 451–62. [Google Scholar]
49. Стиллман Р.А. MORPH-индивидуальная модель для прогнозирования влияния изменений окружающей среды на популяции кормящихся животных. Экологическое моделирование. 2008; 216: 265–76. [Google Scholar]
50. Гимеси А., де Врис П.П., де Бур Т., Нолет Б.А. Уменьшение запасов клубней рдеста фенхеля из-за летнего выпаса водоплавающих птиц. Аква Бот. 2011;94(1):24–8. [Google Scholar]
51. Hidding B, Klaassen M, de Boer T, de Vries PP, Nolet BA. Водное растение демонстрирует гибкое уклонение, спасаясь от нападения лебедей на клубни. Базовая прикладная экол. 2012;13(1):50–8. 10.1016/j.baae.2011.11.002 [CrossRef] [Google Scholar]. Бик-Убберген: SOVON Vogelonderzoek Nederland, 2011 Контракт №: SOVON-отчет о мониторинге 2011/03, Waterdienst-отчет BM10.24.
Водное растение демонстрирует гибкое уклонение, спасаясь от нападения лебедей на клубни. Базовая прикладная экол. 2012;13(1):50–8. 10.1016/j.baae.2011.11.002 [CrossRef] [Google Scholar]. Бик-Убберген: SOVON Vogelonderzoek Nederland, 2011 Контракт №: SOVON-отчет о мониторинге 2011/03, Waterdienst-отчет BM10.24.
53. Шеффер М., Бавеко Дж.М., ДеАнджелис Д.Л., Роуз К.А., ван Нес Э.Х. Супериндивидуумы: простое решение для индивидуального моделирования больших популяций. Экологическое моделирование. 1995; 80: 161–70. [Google Scholar]
54. Холлинг С.С. Некоторые характеристики простых видов хищничества и паразитизма. Канадский энтомолог. 1959; 91: 385–98. [Google Scholar]
55. Эванс М.Е., Кир Дж. Вес и размеры белых лебедей зимой. дичь. 1978; 29: 118–22. [Академия Google]
56. Goss-Custard JD, Caldow RWG, Clarke RT, Durell SEAlVd, Sutherland WJ. Получение параметров популяции на основе индивидуальных вариаций кормодобывающего поведения. I. Эмпирическая модель теории игр распределения куликов-сорок Haematopus ostralegus, питающихся мидиями Mytilus edulis.