Моонзунд
Всего за несколько недель до захвата власти большевиками произошло одно из самых драматичных сражений русского флота
Бой броненосца «Слава» с немецкими кораблями в Моонзундском проливе. Худ. Г.В. Горшков. 1947 год (Фото предоставлено М. Золотаревым)
Строго говоря, это сражение смело можно назвать последней битвой старого, дореволюционного флота. Оно окончилось 4 (17) октября 1917 года – за три недели до того, как перестала существовать прежняя Россия и на ее месте возникла новая страна.
Сила на силу
Моонзундское сражение традиционно рассматривается как часть операции «Альбион», спланированной командованием рейхсхеера (германские сухопутные войска) и кайзерлихмарине (германский военно-морской флот) в начале осени 1917 года. Основная цель операции заключалась в предотвращении атаки с моря на германскую группировку, которая к 19 августа (1 сентября) сумела нанести серьезный удар по русской 12-й армии и два дня спустя взяла Ригу.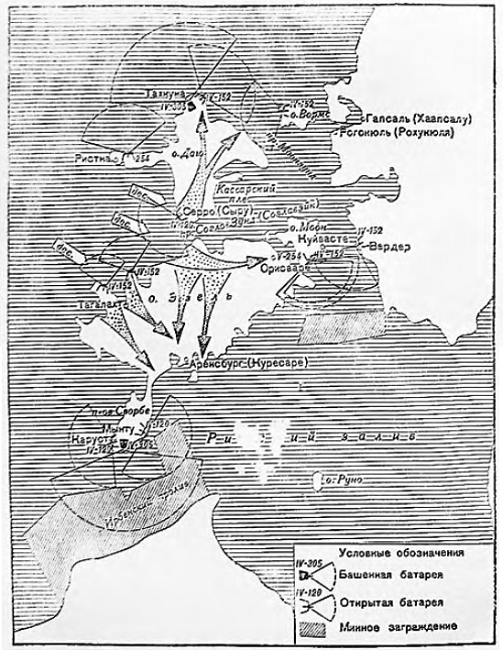
Оборона Моонзундского архипелага. 1917 год
Чтобы добиться такого господства, германское командование выделило значительные силы. Предстояло не только оттеснить русский флот как можно дальше в сторону Петрограда, но и полностью взять под контроль все Моонзундские острова, на которых еще располагался довольно крупный русский гарнизон – две дивизии и несколько батарей береговой артиллерии, в том числе и 305-мм батарея № 43 на мысе Церель (остров Эзель). В своем исследовании «Моонзундская операция Балтийского флота 1917 года», опубликованном в 1928-м, бывший капитан 1-го ранга Алексей Косинский писал: «Из батарей, сооруженных на островах позиции, первое место по своему значению занимала батарея на мысе Церель (№ 43) из четырех 12-дюймовых [305-мм.
Береговая артиллерийская установка 305-мм батареи № 43 на мысе Церель. Остров Эзель (ныне Сааремаа), 1917 год (Фото предоставлено М. Золотаревым)
Куда хуже обстояло дело с кораблями. Даже простое арифметическое соотношение говорит о существенном перевесе немецкого флота: его силы превышали силы русского втрое. С германской стороны насчитывалось около 300 кораблей, включая 9 новейших линкоров типов «Кайзер» и «Кёниг» и линкор «Байерн», 56 эсминцев, 9 легких крейсеров, 6 подводных лодок. Русский флот в Рижском заливе располагал сотней с небольшим кораблей, включая эскадренные броненосцы «Слава» и «Гражданин» (бывший «Цесаревич»), 52 эсминца, 8 подводных лодок, 6 линкоров, 5 броненосных крейсеров и 3 канонерские лодки.
Русский флот в Рижском заливе располагал сотней с небольшим кораблей, включая эскадренные броненосцы «Слава» и «Гражданин» (бывший «Цесаревич»), 52 эсминца, 8 подводных лодок, 6 линкоров, 5 броненосных крейсеров и 3 канонерские лодки.
Ощутимым был и перевес в авиации: германская имела 102 самолета и 6 дирижаблей, а русская – всего четыре десятка аэропланов. Наконец, превосходство наблюдалось и в живой силе. Немцы готовили к десанту 25 тыс. человек при 40 орудиях, 85 минометах и 225 пулеметах, а противостояло им на островах, если не считать береговой артиллерии, около 20 тыс. человек при 60 легких орудиях и 140 пулеметах.
Но главным преимуществом немецкого Морского отряда особого назначения, сформированного для захвата Моонзундского архипелага, являлась дисциплина. Русским войскам нечего было ей противопоставить. Развернутая большевистскими представителями в армии и на флоте антивоенная пропаганда привела к полному развалу вооруженных сил. Приказы выборных командиров обсуждались до тех пор, пока не теряли всякий смысл.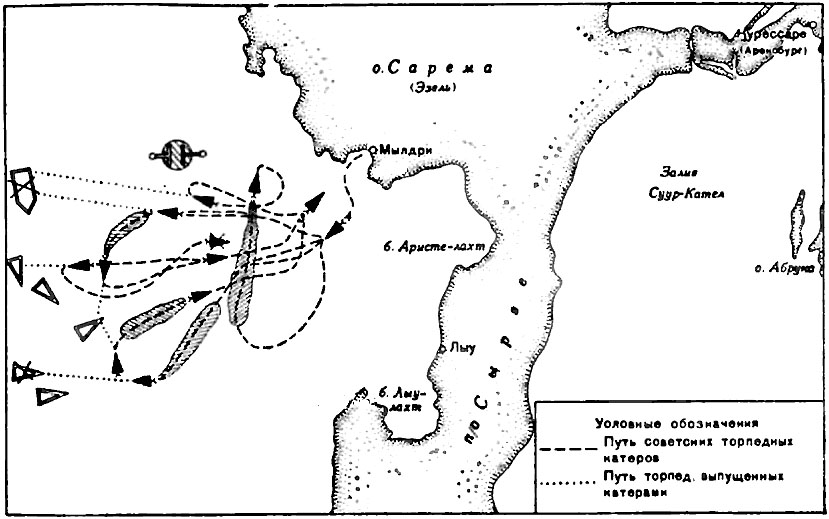 Ярчайшим примером служит отказ экипажа минного заградителя «Припять» выполнить распоряжение командующего Морскими силами Рижского залива вице-адмирала Михаила Бахирева о постановке мин в проливе Соэлозунд – месте, через которое началось морское наступление в рамках операции «Альбион»…
Ярчайшим примером служит отказ экипажа минного заградителя «Припять» выполнить распоряжение командующего Морскими силами Рижского залива вице-адмирала Михаила Бахирева о постановке мин в проливе Соэлозунд – месте, через которое началось морское наступление в рамках операции «Альбион»…
Атака на Моонзунд
С учетом всего этого становится понятно, почему первый удар, который 29 сентября (12 октября) 1917 года немецкий особый отряд нанес по острову Эзель, почти сразу увенчался успехом. Дезорганизованные русские сухопутные войска практически не сопротивлялись. Стойкость проявили лишь отдельные подразделения, например Ревельский морской батальон смерти капитана 2-го ранга Павла Шишко, который несколько суток сдерживал натиск немцев на Ориссарской дамбе, соединяющей острова Эзель и Моон. Однако их усилий не хватало, чтобы сорвать вражеское наступление. Тем более что рядом целые роты сдавались в плен: зачинщики насильно волочили за собой тех, кто отказывался поднять руки.
Неудивительно, что генерал фон Чишвиц впоследствии подчеркивал: «Общие потери [германской группировки. – А. Т.], считая и флот, исчисляются в 400 чел., что составляет каплю в море по сравнению с 20 000 русских, взятых в плен…»
Между тем сопротивление на море оказалось гораздо сильнее, чем на суше. Именно морская часть операции «Альбион», которую и называют собственно Моонзундским сражением, пошла далеко не так гладко, как рассчитывали германские генштабисты.
Первый этап этого сражения развернулся на входе в пролив Соэлозунд, севернее острова Эзель, где в течение 30 сентября (13 октября) шла корабельная дуэль. Только на следующий день корабли вошли в боевое соприкосновение на Кассарском плесе. Для русских оно закончилось потерей эскадренного миноносца «Гром» – одного из эсминцев типа «Новик», который попал под залп германского линкора «Кайзер».
Много позже, когда Моонзундское сражение стало предметом советской героизации, эсминец «Гром» в буквальном смысле слова подняли на щит: родилась красивая легенда о минном старшине Федоре Самончуке, который в одиночку торпедировал приближавшийся к подбитому кораблю германский эсминец, а потом бросил в крюйт-камеру факел и взорвал «Гром», чтобы тот не достался врагу.
Однако эту версию совершенно не подтверждает отчет вице-адмирала Михаила Бахирева, командовавшего русскими кораблями в ходе Моонзундского сражения. Вот что он писал об этом эпизоде: «Доблестный командир «Храброго» старший лейтенант Ренненкампф подошел кормой к носу «Грома» и в 14 часов 55 минут принял с него команду. В это время неприятель весь свой огонь сосредоточил по этим двум кораблям. Расчет командира «Храброго» был в том, чтобы подойти к «Грому», стравить к себе на палубу его якорь и таким образом продолжать буксирование. На «Громе» было большое смятение, и, не дождавшись подхода канонерской лодки, человек 15 бросились в воду; конечно, подбирать их не было времени, и с «Храброго» бросили им спасательные круги и тузик. Командир «Грома» не хотел оставлять своего корабля, и его на лодку перетащили силой.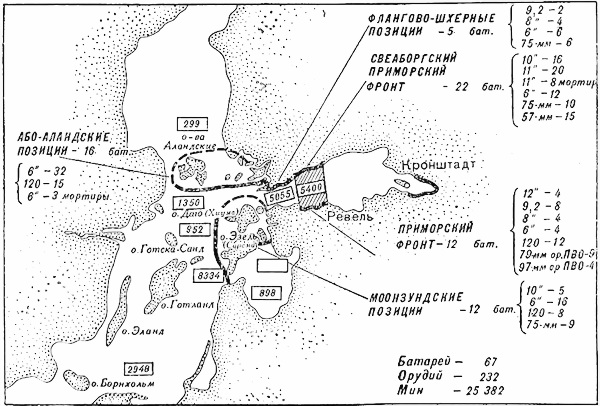
Последний бой «Славы»
На следующий день, 2 (15) октября, произошло еще одно важное событие, приблизившее развязку Моонзундского сражения: пала самая сильная из береговых батарей Эзеля – № 43. Алексей Косинский свидетельствовал: «Неприятельские корабли подошли к Церелю в тумане, видимо, сами не ожидая такой близости к батарее, так как даже орудия на них были повернуты по-походному, что дало батарее время приготовиться к бою. Однако после второго нашего залпа неприятель открыл огонь из своих орудий. Бой продолжался с небольшим перерывом около часа. <…> Несмотря на медленность и неудовлетворительность нашей стрельбы, на трудность управления ею из-за разбрасывания снарядов, неисправности приборов и выхода из действия одного за другим трех орудий, все же в начале боя батарее, по-видимому, удалось вывести из строя один дредноут. В то же время неприятель, имея на своей стороне такие преимущества, как большое число орудий, дисциплинированный огонь, кучность падений снарядов, быстрота стрельбы (промежутки между залпами – 30–40 секунд, тогда как наши двухорудийные залпы давались через 2 минуты), не имел ни одного попадания, причем ближайший снаряд лег от батареи в 30 саженях».
Алексей Косинский свидетельствовал: «Неприятельские корабли подошли к Церелю в тумане, видимо, сами не ожидая такой близости к батарее, так как даже орудия на них были повернуты по-походному, что дало батарее время приготовиться к бою. Однако после второго нашего залпа неприятель открыл огонь из своих орудий. Бой продолжался с небольшим перерывом около часа. <…> Несмотря на медленность и неудовлетворительность нашей стрельбы, на трудность управления ею из-за разбрасывания снарядов, неисправности приборов и выхода из действия одного за другим трех орудий, все же в начале боя батарее, по-видимому, удалось вывести из строя один дредноут. В то же время неприятель, имея на своей стороне такие преимущества, как большое число орудий, дисциплинированный огонь, кучность падений снарядов, быстрота стрельбы (промежутки между залпами – 30–40 секунд, тогда как наши двухорудийные залпы давались через 2 минуты), не имел ни одного попадания, причем ближайший снаряд лег от батареи в 30 саженях».
Михаил Бахирев в годы командования эсминцем «Амурец». 1908–1910 годы (Фото предоставлено М. Золотаревым)
Но затем случилось непредвиденное. В отчете Михаила Бахирева читаем: «Вчера (1 октября) при стрельбе по 305-мм батарее № 43 неприятельских кораблей от северо-запада прислуга двух орудий разбежалась, у третьего орудия осталась половина ее и неприятелю отвечали только два (вернее, полтора) орудия. Повреждений, убитых и раненых нет. Капитан 2-го ранга Кира-Динжан передал мне, что в ночь на 2 октября б Иду [на] Куйваст»».
А через два дня после потери важного узла обороны разыгрался последний акт Моонзундского сражения – бой на рейде поселка Куйвасту в проливе Моонзунд. Днем ранее в Рижский залив с соответствующим охранением вошли линкоры «Кёниг», «Кронпринц» и три легких крейсера, которые предполагали отсюда через Кассарский плес и Моонзундский пролив выйти в Финский залив. Германское командование считало, что заманило русских в ловушку, но наши моряки сумели доказать, что это не так.
Утром 4 (17) октября капитан находившегося в дозоре эсминца «Деятельный» по радио известил Бахирева о том, что противник направляется к рейду Куйвасту. Навстречу врагу двинулись флагман – крейсер «Баян» – и эскадренные броненосцы «Слава» и «Гражданин». Силы были явно неравны, но Бахирев надеялся на прикрытие минного заграждения и огонь береговых батарей: в такой позиции обороняться можно было весьма успешно. И тем не менее командующий приказал подготовить к затоплению в проливе (точнее, в прорытом русскими моряками канале, который позволял крупным боевым кораблям проходить по проливу) угольные транспорты «Глагол» и «Покой».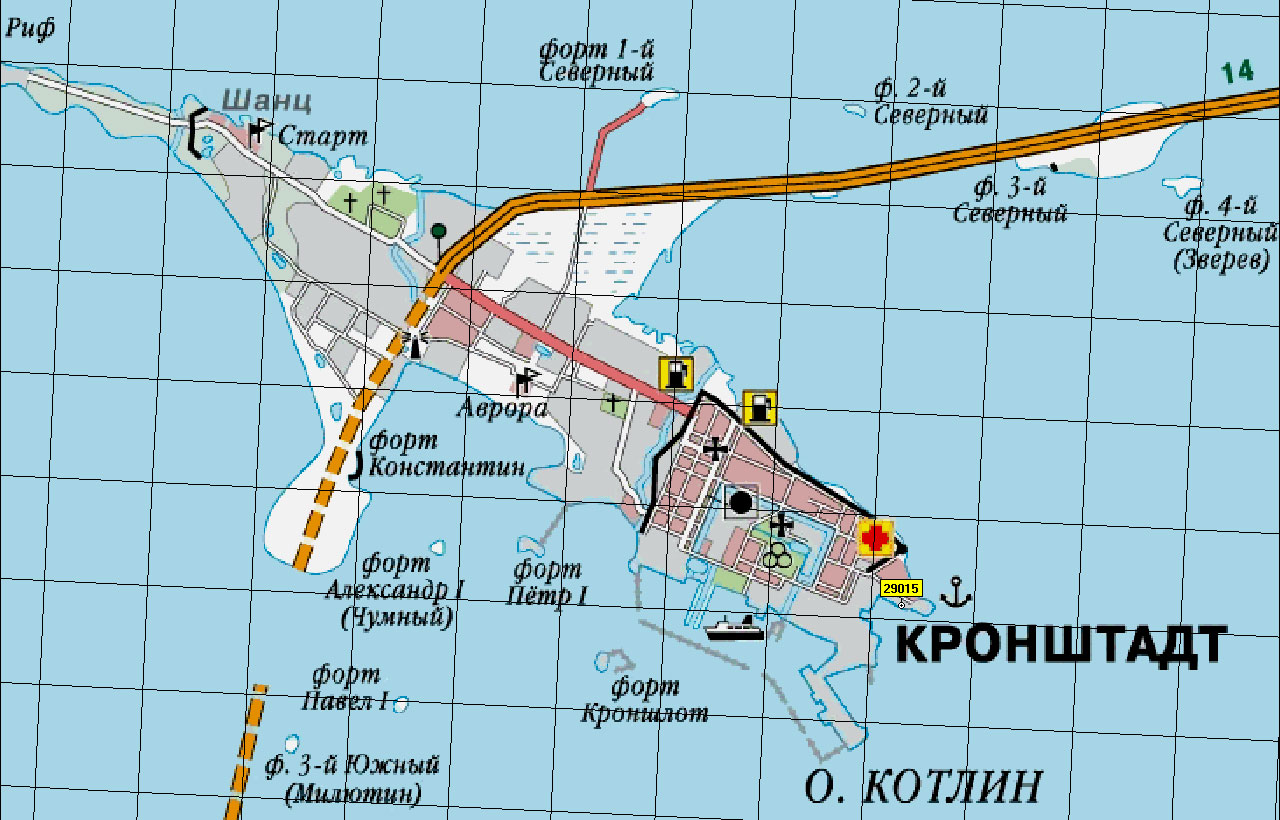
Бой начался с выстрелов береговой батареи острова Моон по германским тральщикам, пытавшимся преодолеть минную банку. Батарейный огонь поддержала «Слава», накрыв врага с предельной дистанции из носовых орудий, и противник отошел. В перестрелку вступили «Кёниг» и «Кронпринц», но их снаряды ложились с большим недолетом. Так что после того, как «Слава» накрыла еще один миноносец, неприятель поспешил разорвать огневой контакт, напоследок дав несколько залпов по батареям на Мооне. В 11:20 на «Баяне» подняли сигнал: «Полубригаде линейных кораблей адмирал изъявляет свое удовольствие за отличную стрельбу».
Правда, этот успех дорого обошелся «Славе». Получив передышку, ее комендоры осмотрели носовую 305-мм башню и поняли, что она к дальнейшей стрельбе непригодна: на обоих орудиях нельзя было открыть замки, поскольку сломались вдвигавшие их бронзовые шестеренки зубчатки и опустились рамы. Таким образом, эффективно действовать против двадцати 305-мм немецких пушек теперь могли лишь два орудия кормовой башни «Славы». Орудия «Гражданина» были менее дальнобойными, а из всех орудий батареи на Мооне исправными остались только два, да и те очень медлительные.
Орудия «Гражданина» были менее дальнобойными, а из всех орудий батареи на Мооне исправными остались только два, да и те очень медлительные.
И тогда Бахирев просигналил отступление: русские корабли начали отходить на север. Увидев это, неприятель бросился вдогонку, и тут в полной мере проявилось превосходство в массе совокупного залпа и моральной силе. Быстро шедшая в авангарде «Слава» получила семь попаданий, причем все – перелетами через «Гражданина». Три из них пришлись ниже броневого пояса: корабль сразу принял почти 1,5 тыс. тонн воды и сильно осел на левый борт. Чтобы выровнять его, команда открыла кингстоны правого борта и затопила коридоры. При этом осадка увеличилась настолько, что об отступлении «Славы» через Моонзунд уже не могло быть и речи…
Бахирев писал в отчете: «В 13 часов, когда на расстоянии 1/2–3/4 мили от входа в глубоководный Моонзундский канал «Баян» обходил «Гражданина» и «Славу», командир последнего доложил мне о бедственном состоянии корабля и просил разрешения взорвать его. С просьбой о помощи раздались несколько истеричных голосов с верхней палубы. Разрешение взорвать корабль мной было дано командиру «Славы», и я приказал ему, пропустив вперед «Баян» и «Гражданин», затопить корабль в самом канале – при входе в него. На всякий случай напомнил командиру – о необходимости уничтожения секретных карт, книг и документов. В 13 часов 10 минут с нагонявшего отряд «Дельного» [эскадренный миноносец. – А. Т.], наблюдавшего за неприятелем, был получен семафор: «Моонские батареи больше не работают». В 13 часов 15 минут «Баян» и вслед за ним «Гражданин» вошли в канал и малым ходом пошли по нему на север». А три четверти часа спустя прогремел взрыв: сработали мины, установленные в артиллерийских погребах «Славы».
С просьбой о помощи раздались несколько истеричных голосов с верхней палубы. Разрешение взорвать корабль мной было дано командиру «Славы», и я приказал ему, пропустив вперед «Баян» и «Гражданин», затопить корабль в самом канале – при входе в него. На всякий случай напомнил командиру – о необходимости уничтожения секретных карт, книг и документов. В 13 часов 10 минут с нагонявшего отряд «Дельного» [эскадренный миноносец. – А. Т.], наблюдавшего за неприятелем, был получен семафор: «Моонские батареи больше не работают». В 13 часов 15 минут «Баян» и вслед за ним «Гражданин» вошли в канал и малым ходом пошли по нему на север». А три четверти часа спустя прогремел взрыв: сработали мины, установленные в артиллерийских погребах «Славы».
Лежащий на грунте взорванный броненосец «Слава». Моонзундский канал, конец 1917 года (Фото предоставлено М. Золотаревым)
«Не до ордена – была бы Родина»
Формально Моонзундская операция увенчалась победой германских войск: им удалось полностью захватить архипелаг.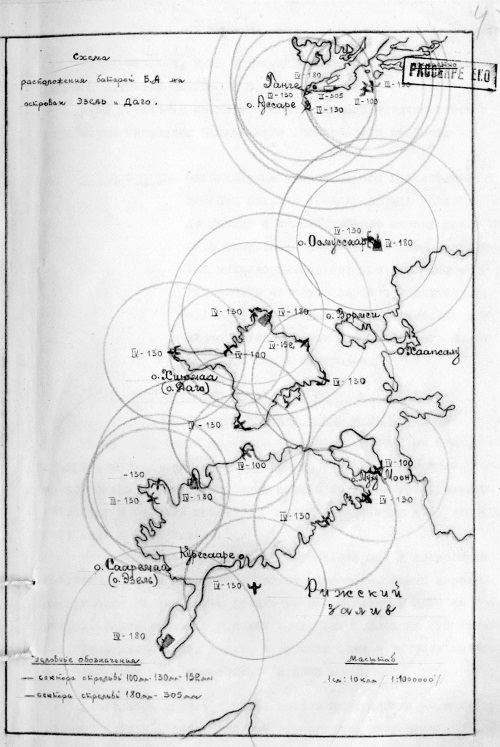 При этом девять немецких кораблей были потоплены, еще больше оказались повреждены, тогда как русская эскадра безвозвратно потеряла всего два судна – «Гром» и «Славу». Число погибших было примерно одинаковым – не более 400 человек с каждой стороны, но русская армия потеряла свыше 20 тыс. человек пленными. Зато затопленные на входе в Моонзундский канал «Слава» и угольные транспорты все-таки закрыли германской флотилии путь к Кронштадту и Петербургу.
При этом девять немецких кораблей были потоплены, еще больше оказались повреждены, тогда как русская эскадра безвозвратно потеряла всего два судна – «Гром» и «Славу». Число погибших было примерно одинаковым – не более 400 человек с каждой стороны, но русская армия потеряла свыше 20 тыс. человек пленными. Зато затопленные на входе в Моонзундский канал «Слава» и угольные транспорты все-таки закрыли германской флотилии путь к Кронштадту и Петербургу.
Впрочем, все эти выводы делались уже много лет спустя. А тогда, через три недели после сражения, грянула Октябрьская революция и ни русским, ни германским военным некогда было анализировать недавнюю битву. Позднее каждая из сторон представляла исход Моонзундского сражения как свой успех. Однако, рассуждая здраво, можно с уверенностью сказать: русский флот в своем последнем крупном сражении одержал если не безусловную победу, то по очкам – наверняка.
И победа эта тем более ценна, что добились ее наши моряки вопреки очень и очень многим обстоятельствам.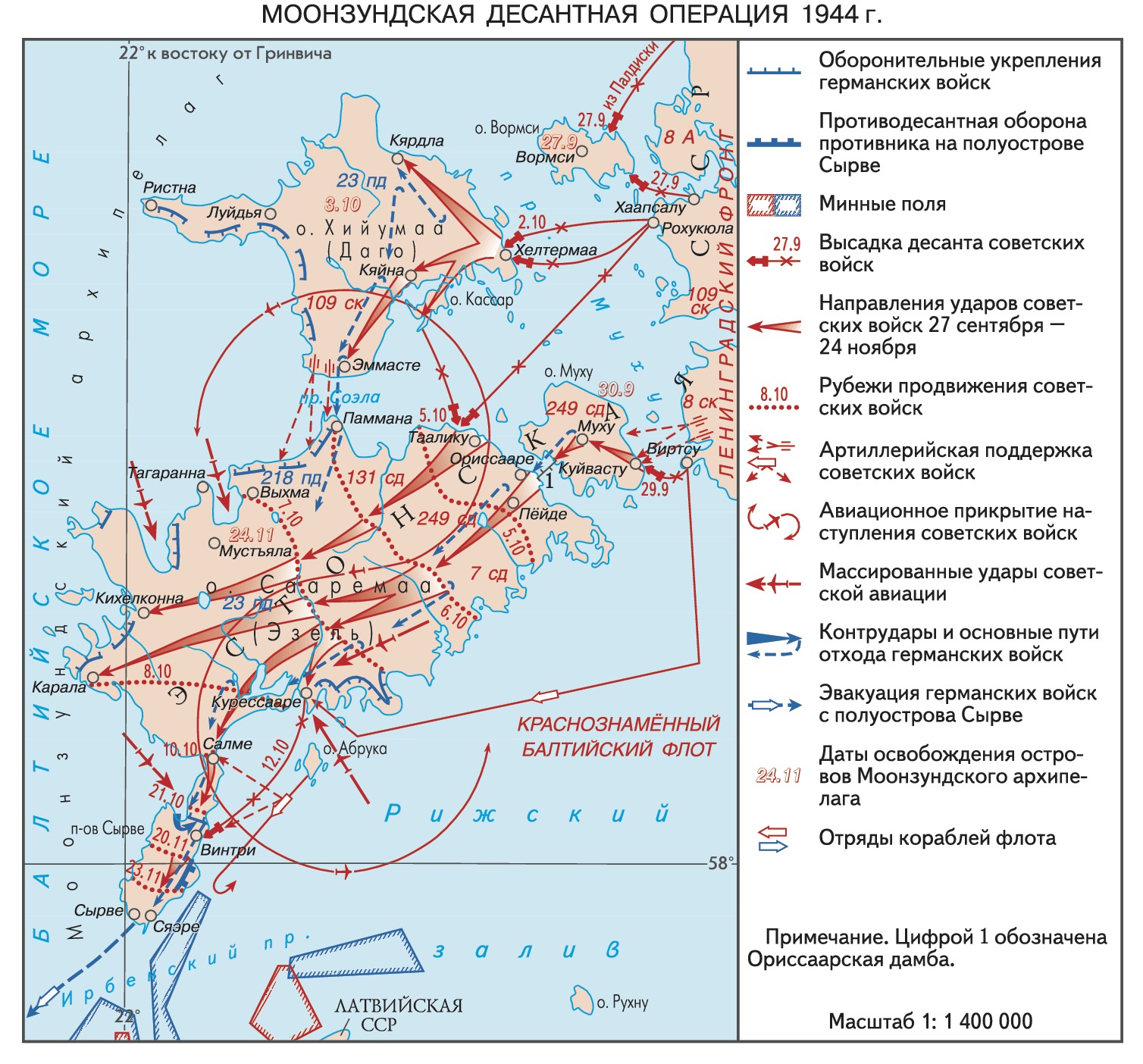 Коллапс системы военного управления, беспрецедентное падение флотской дисциплины, отсутствие координации между подразделениями и видами войск – все это могло свести на нет любые усилия адмирала Бахирева и его подчиненных. Но – не свело.
Коллапс системы военного управления, беспрецедентное падение флотской дисциплины, отсутствие координации между подразделениями и видами войск – все это могло свести на нет любые усилия адмирала Бахирева и его подчиненных. Но – не свело.
Антон Трофимов
Батарея № 43 (Церель)
Батарея № 43 — крупнокалиберная береговая батарея в составе Моонзундской позиции Морской крепости Императора Петра Великого, защищавшая Ирбенский пролив. Находилась у мыса Церель
1. История
Строительство под руководством Н. И. Унгермана началось осенью 1916 года и было закончено в апреле 1917 года.
В сентябре 1917 года немцы, стремившиеся прорваться в Рижский залив, начали интенсивно бомбить батарею, орудия которой не позволяли производить минное траление в Ирбенском проливе. 18 сентября в результате одного из налётов произошёл взрыв в пороховом погребе, в результате чего 121 человек погиб. Командир батареи был ранен.
2. Описание
Батарея № 43 имела ряд серьёзных недостатков: установки стояли открыто, без какой бы то ни было броневой защиты. Бетонные брустверы начали строить перед самым началом операции «Альбион». И кроме того, батарею в ущерб маскировке слишком далеко продвинули на юг, к проливу. Это, конечно, повлияло на исход Моонзундского сражения, но это влияние не было решающим. Крайне неудачным оказалось устройство погребов. Открытые с тыла, они имели здесь широкие входы для тележек с боеприпасами. Входы закрывались лишь дубовыми дверями, и это обстоятельство имело трагические последствия…
3. Образ в искусстве
Образ в искусстве
Связанные с батареей № 43 события 1917 года нашли отражение в романе В. С. Пикуля «Моонзунд» и одноимённом художественном фильме. Командир батареи старший лейтенант Николай Бартенев выведен в романе под именем старшего лейтенанта Сергея Артеньева. Одноимённый фильм снимался на 3-м форту Лиепаи Кароста, при этом вместо 305/52-мм орудий использовалось одно 210-мм орудие Бр-17 и деревянные макеты.
Дата публикации:
05-16-2020
Дата последнего обновления:
05-16-2020Береговая батарея № 43. Церельская батарея.
Батарея № 43. Церельская батарея. 4 орудия калибром 305 мм (12 дюймов). Открытая стационарная. Расположена рядом с маяком Сааре. Хорошо сохранились все четыре основания орудий.
Расположена рядом с маяком Сааре. Хорошо сохранились все четыре основания орудий.
Самая мощная батарея на мысе Церель. Командир батареи старший лейтенант Бартенев. Церельскую батарею строил известный русский фортификатор Унгерман. Вступила в строй в апреле 1917 года. Состояла из четырех открытых позиций 305-мм орудий и двух бронированных капониров. Между пристанью Мынту и батареей была проложена 4,5 километровая узкоколейная (колея 750 мм) железнодорожная ветка. На линии использовался паровоз типа 0-3-0 фирмы «Оренштейн и Коппель» мощностью 90 л.с., изготовленный в Германии в 1914 году. Восьмитонные платформы были сделаны в 1913 году на вагонном заводе «Артур Коппель» в Санкт-Петербурге.
В 1907 году для линкоров типа «Севастополь» была спроектирована 305/52-мм пушка. Заказ на изготовление пушек получил Обуховский сталелитейный завод (ОСЗ) в Санкт-Петербурге. И началу 1917 года Морское ведомство получило 126 орудий, которыми были вооружены балтийские и черноморские линкоры. В 1910 году Главное артиллерийское управление заказало ОСЗ 16 береговых установок 305/52-мм пушек затем последовали еще заказы, и к сентябрю 1917 года было изготовленно 36 установок. Лафеты 305/52-мм орудий изготавливал Металлический завод в Санкт-Петербурге.
В 1910 году Главное артиллерийское управление заказало ОСЗ 16 береговых установок 305/52-мм пушек затем последовали еще заказы, и к сентябрю 1917 года было изготовленно 36 установок. Лафеты 305/52-мм орудий изготавливал Металлический завод в Санкт-Петербурге.
На Балтике береговые батареи с 305/52-мм орудиями были построены на островах Эре, Эзель и Даго, а также в крепостях — форт Ино и форт Алек- сеевский (Красная горка). Планировалось строительство такой же батареи в Усть-Двинске (Рига). Паровоз ((0ренштейн и Береговая установка 305/52-мм пушки представляла собой внушительное сооружение. Ствол пушки имел длину 16 метров и весил более 50 тонн. Высота установки 6 метров, общий вес 120 тонн. Несмотря на огромные размеры заряжение орудия происходило вручную. Снаряд весом 470 кг поднимался на линию подачи ручной лебедкой и далее 6 человек прислуги прибойником досылали его в ствол. Также вручную досылались и пороховые заряды весом 132 кг. Фугасный снаряд образца 1911 года нёс 60 кг взрывчатого вещества, имел начальную скорость 800 м/сек и дальность полета 28 км.
На каждой пушке имелся свой перископический прицел Цейса. Общее управление огнем батареи осуществлялось приборами управления стрельбой системы Гейсмера с 1,5 — метровым дальномером.
Для обороны Ирбенского пролива русский флот за три года войны выставил около 10000 мин. В 1917 году, в связи с захватом немцами Курляндского берега, русские выставили большое минное заграждение у мыса Домеснес (Колкасрагс).
Немецкий флот неоднократно пытался проводить траление мин в Ирбенс- ком проливе, но каждая попытка тралить фарватер отражалась огнем батарей Цереля. Немцы понимали, что не уничтожив 305-мм батарею, они не смогут осуществить прорыв крупными силами в Рижский залив.
Для германских линейных кораблей 43-я батарея была целью номер один. После постоянных обстрелов батарея отвечала огнем двух орудий, так как прислуга двух других разбежалась. Позиции батареи подвергались бомбардировкам с воздуха. После очередной воздушной атаки 17 сентября 1917 года загорелся склад пороховых зарядов. Строительство батареи не было закончено, склад пороховых зарядов и склад снарядов находились в бревенчатых погребах, стена между склаццми прогорела и последовал взрыв сорока четырех 305-мм снарядов. В результате взрыва выбыло из строя 7 офицеров и 114 матросов. К середине октября кайзеровские войска подошли к батарее с суши. Орудия батареи не имели кругового сектора обстрела и не могли противодействовать войскам, наступавшим на мыс Церель по полуострову Сворбе. Немцы предъявили батарее ультиматум о сдаче. Батарейцы ответили: «Русские в плен не сдаются. Долг повелевает нам драться до последнего снаряда».
43-я батарея держалась до 15(2) октября. Под огнем немецких линкоров личный состав покинул батарею. На батарее остались 20 человек во главе с командиром, с намерением взорвать позиции и орудия. Попытки взорвать двенадцатидюймовые орудия оказались тщетными. Подрывная команда и командир батареи попали в немецкий плен, откуда возвратились только в 1919 году. Судьба 43-ей батареи вообще связана с цепью трагических обстоятельств. Не имея сведений о том, в чьих руках находится батарея, русское командование послало линейный корабль «Гражданин» («Цесаревич») с приказом обстрелять и уничтожить батарею. Линкор дал несколько залпов, но с наступлением темноты стрельбу прекратил.
Командир 43-ей батареи — старший лейтенант Бартенев стал прообразом главного героя романа Валентина Пикуля «Моонзунд» — старшего лейтенанта Артемьева. Всё действие романа подводило русского офицера к точке наивысшего напряжения событий — обороне мыса Церель, где решалась судьба Российской империи. С потерей батарей Цереля Прибалтика была потеряна для России на долгие десятилетия.
Путешествия по Сааремаа. Сегодня маяк Сырве и пляж
Время, отведенное на посещение острова Сааремаа, неумолимо подходило к концу, а отпуск к своему экватору. Мы уже посмотрели на многие достопримечательности острова и в плане остался только маяк Сырве. Туда и отправимся, а заодно заедем на пляж (какой именно – читаем в статье) и к монументу защитникам острова.
Чем интересен маяк Сырве и почему он стоит того, чтобы его посетить
Каждая достопримечательность обладает неким качеством, которое выделяет её среди похожих мест или сооружений и это качество обычно обозначается прилагательным “самый” – самый высокий, самый древний, в общем самый-самый. Маяки – отдельная тема. Мне самому нравятся эти сооружения, но что именно меня в них притягивает точно сказать не могу. Маяк, по своей сути, исключительно техническое сооружение. Не вызывают же такого интереса, например, мачты городского освещения или вышки сотовой связи. Маяки интерес вызывают и притягивают к себе не просто туристов, есть целые сообщества любителей, ценителей и знатоков маяков в разных частях света. Возможно, все дело в том, что мы видим в них нечто противоположное – надежду и одиночество одновременно. Как правило, маяки возводятся на мысах, вдали от цивилизации и их смотрители ведут одинокий, иногда почти отшельнический образ жизни. Для морских путников маяк – надежда и ориентир, иногда спаситель.
За время пребывания на Саарема мы уже познакомились с малоизвестным маяком на полуострове Кюбассааре, а еще раньше мы побывали у разных маяков в России и Латвии.
Ирбенский пролив считается весьма сложным для навигации местом. Оконечность мыса Сырве и латвийский берег отделяют всего 30 км. Место это для судоходства сколь важное, столь и опасное. Даже не будучи военным стратегом, глядя на карту, на мыс Сырве почти упирающегося своим острием в мыс Колка, что расположен материковой латвийской уже стороне, понимаешь, что Ирбенский пролив должен был (и был) стратегически важным местом в любой войне, так как образует самый широкий вход из Балтики в Рижский залив. Лишь немного окунувшись в историю двух последних мировых войн узнаешь, что интуиция не подвела.
Интересный, хотя и немного мрачноватый пейзаж в обеих войнах не единожды становился поводом для жестоких и кровопролитных боёв за возможность занять стратегически важную точку, которая позволяла контролировать вход в гавань. Неудивительно, что в 1944 году каменная башня маяка была уничтожена. Но маяк имел крайне важное значение для мореходства в этом районе и наскоро в 1949 году была сооружена деревянная башня. Однако возросший поток судов в проливе требовал обеспечения безопасности и, как уже было сказано, в 1960 году появился маяк в его нынешнем виде. По тем временам маяк имел современную светооптическую аппаратуру с белым проблесковым огнем, что обеспечивало видимость маяка для судов на расстоянии 19 морских миль, в прилегающем здании также был установлен радиомаяк с радиусом действия в 180 км.
А 70-80-х годах прошлого века здесь использовался весьма редкий вид маяков – плавучий маяк. Сейчас это весьма редкое и уникальное в общем-то судно используется в качестве музея и базируется в Калининграде. Вот так выглядит это необычное судно:
Для тех, кто интересуется морской тематикой, думаю, небезинтересно будет посмотреть короткий фильм еще 80-х годов прошлого века о плавучем маяке.
Об опасностях, которые таит в себе Ирбенский пролив говорит и то, что о нем бывалые моряки слагали легенды. Вот одна из них:
Согласно легенде образовался такой узкий пролив, когда дьявол решил построить мост между Сааремаа и материком, но не успел до захода солнца.
Вернемся, однако на мыс к маяк Сырве (Sõrve), самому старому или во всяком случае одному из самых старых. Его создание относится к 1646 году. Глядя на это сооружение, конечно, возникают законные сомнения в том, что в середине XVII века строили такие маяки. Сомнение совершенно верное. 350 лет назад маяк выглядел совершенно иначе, ну точнее мы почти наверняка знаем, что он выглядел иначе, так как нынешний его облик относится к совсем недалекому 1960 году.
А тогда в 1646 году, когда на месте Эстонии правил еще Ливонский орден, некий купец по имени Эберт-Деллингсгаузен построил маяк, чтобы безопасно возить свои товары через Ирбенский пролив. Кстати, название Сырве (по названию мыса, на котором он и стоит) маяк получил значительно позднее, а первое его название – Церель, или Свальферорт. Тогда это была деревянная башня высотой 34,7 метра. В 1807 году, когда территория стала частью Российской империи и именовалась Лифля́ндской губе́рнией высота маяка была увеличена, а место деревянной башни возведена каменная.
Разумеется, об электроэнергии тогда никто не слыхивал и маяк Сырве, как впрочем и все другие маяки, освещался кострами, а топливом служили дрова, смола и минеральные масла.В середине XX века была построена железо-бетоная башня. Сейчас маяк Сырве остается действующим, а в прилегающем здании для туристов устроен музей эстонских маяков и симпатичное кафе.
Военное кладбище и монумент у Техумарди
По дороге к маяку или на обратном пути (кому как удобнее) можно заехать в местечко Техумарди.
Адрес: Tehumardi, Salme, Saaremaa,
GPS координаты: 58.176881, 22.254504 58° 10′ 36.77″, 22° 15′ 16.21″
Монумент расположен на бывшем поле боя, где сражались воины 917-го, 925-го стрелковых полков и передового отряда 249-й дивизии Эстонского корпуса во Второй Мировой Войне. Бой у Техумарди, происшедший во время Моонзундской десантной операции в 1944 году, был одним из самых кровопролитных сражений за освобождение острова Сааремаа.
Монумент представляет собой мощный, вздымающийся на 21 метр ввысь стилизованный меч, рукоять которого упирается в землю, лезвие направлено вверх, символизируя опасность войны. Крестовина меча находится в нескольких метрах от поверхности земли.
В доломитных блоках, расположенных на концах крестовины меча, высечены рельефные лица воинов. Рядом расположено братское кладбище, где установлены 90 ромбовых блоков с фамилиями погибших воинов. Мемориал открыт в 1966-ом году , авторы- Riho Kulla, Matti Variku ja Allan Murdmaa.
Купальный пляж Мяндъяла (Mandjala beach)
Добраться до пляжа совершенно не сложно, к нему ведет асфальтированная дорога.
Адрес: Эстония, 93871 Saare maakond, Эстония
Координаты GPS: 58.211958, 22.323011
В сезон и особенно в выходные дни народу здесь может быть достаточно много, эстонцы любят море и любят отдыхать на своих курортах благо ехать до них совсем недалеко.
Где остановиться рядом с пляжем и Техумарди
Сааремаа славится шикарными СПА-отелями и гостевыми домами в экологически чистых местах. Одним из таких гостевых домов является Järve Cottages. Эстонцы обожают такой эко-деревенский стиль.
Но при внешней аскетичности, все удобства в таком домике будут присутствовать, здесь иначе не бывает.
Так завершилось наше почти недельное пребывание на острове Саамремаа, о котором у нас останутся исключительно положительные воспоминания. Далее мы отправляемся снова к Балтийскому морю, но уже литовскому. Хотелось сказать в соседнюю Литву, но глянув на карту, понимаем, что придется сначала проехать почти через всю Латвию.
Поделиться ссылкой:
Статья обновлена:7/04/20
Понравилось это:
Нравится Загрузка…
Оборона Моонзунда 1941 год | Самое интересное в истории Российского флота
Маяк Тахкуна
Фото с сайта Discoveric
С 6 сентября по 22 октября 1941 года — оборона Моонзундских островов — одна из самых героических и трагических страниц Великой Отечественной войны. Находясь в сотнях километров западнее линии фронта, защитники архипелага своей стойкостью, мужеством и самоотверженной обороной сковали крупную группировку сил противника. С этих островов наша морская авиация нанесла первые бомбовые удары по Берлину. Этот коллективный подвиг на далеких от эпицентра сражений островах помог сорвать планы гитлеровцев по захвату Ленинграда и уничтожению Балтийского флота.
Оборона продолжалась до полного исчерпания всех возможностей сопротивления врагу. Последний бой за Моонзунд произошел на мысе Тахкуна. Матрос Николай Чиж вел его на верхней площадке маяка, а когда кончились патроны, он раскинул руки и бросился вниз, на острые камни… Позднее эти острова заслуженно называли Балтийским Брестом. Герои Моонзунда с честью выполнили свою боевую задачу, однако в те времена их подвиг не получил признания. На долгие годы было предано забвению и имя их командующего генерал-лейтенанта А.Б. Елисеева.
Подготовка
Моонзундский архипелаг включает три крупных и множество мелких островков, которые простираются с севера на юг всего на 150 километров. В далеком прошлом на них укрывались морские пираты, и с тех пор не было на Балтике таких военных конфликтов, которые обошли бы стороной эту землю. И это понятно – три небольших острова позволяют надежно перекрыть входы в Рижский и Финский заливы, полностью контролировать обстановку в центральной и восточной части Балтийского моря. Это значит прикрыть многие жизненно важные центры северо-западной части нашей страны.
Именно здесь, на Моонзунде и в Рижском заливе, развернулись самые ожесточенные бои между русским и германским флотами в годы Первой мировой войны. Тогда в 1918 году II съезд военных моряков Балтики постановил: «Матросы Балтики докажут всему миру, что революционный флот, защищая революционную Россию, погибнет, но не отступит перед флотом германского империализма» (смотрите статью «Эсминец «Гром» — последний бой»). И 23 года спустя, принимая свой последний бой на полуострове Тахкуна, моряки вновь поклялись доказать всему миру, что они умеют умирать с честью, выполнив свой долг перед Родиной.
К этой войне усиленно готовились. На островах Моонзунда спешно достраивали береговые батареи, соорудили более 250 дотов и дзотов, прикрывая их минными полями и колючей проволокой. Довольно мощная береговая артиллерия архипелага, включавшая 213 орудий, готовилась к отражению противника со стороны моря и была плохо приспособлена к стрельбе по берегу. Гарнизон островов насчитывал тогда около 24 тысяч человек. Ему были приданы 12 истребителей и 6 торпедных катеров. Учитывая большую протяженность береговой линии, этого было совершенно недостаточно.
Начало боевых действий
С началом войны на базе воинских частей и оборонительных сооружений Моонзундских островов создали соединение Береговой обороны Балтийского района (БОБР) под командованием генерал-майора (с 16.09.1941 г. генерал-лейтенанта) А.Б. Елисеева. Вся ответственность за оборону БОБР и Рижского залива полностью возлагалась на Балтийский флот. Фактически это была борьба за обеспечение фланга огромного фронта, упиравшегося в Балтийское море.
Карта Моонзундского архипелага
Уже в середине июля 1941 года противник оказался в непосредственной близости от островов. Тогда Военный совет флота приказал коменданту островного района генерал-майору А.Б. Елисееву 18 июля высадить на материк десант и отбросить немцев к Пярну. Эта задача была выполнена отлично, и противник почти два месяца даже не пытался захватить острова. В начале августа с острова Сааремаа наша морская авиация нанесла первые удары по Берлину. Налеты продолжались до 4 сентября, зарево пожаров в Берлине было видно далеко за пределами города.
Большие потери, вызванные активной обороной наших войск, резко снизили темпы наступления фашистских армий. У них возникла острая потребность в подкреплениях, технике и боеприпасах. Тогда немецкое командование решило использовать порты Рижского залива для снабжения наступавшей на Ленинград группы армий «Север». Наши корабли и морская авиация, базировавшиеся на Моонзунде, а также береговые батареи острова Сааремаа серьезно нарушали морские перевозки в этом районе. Здесь 6 июля произошел первый артиллерийский бой надводных кораблей.
Бой кораблей КБФ в Ирбенском проливе
C картины А.А. Ефимова
После захвата Таллина немецкое командование начало спешно готовиться к штурму архипелага. Оно выделило крупные силы пехоты, обеспечив им мощную поддержку с моря и с воздуха. Утром 8 сентября после сильной артподготовки немецкие войска высадились на остров Вормси. Гарнизон, состоявший из двух неполных рот, сражался мужественно. Только в районе маяка фашисты оставили около 300 трупов. Однако противнику удалось захватить основные пункты нашей обороны. Защитники острова вели бой до последней возможности, и лишь немногим из них удалось уйти на Хийумаа.
Упорные бои за каждую позицию, за каждый остров
К сентябрю немцы сосредоточили на побережье группировку общей численностью до 50 тысяч человек и около 350 высадочных средств. Ее поддерживали бомбардировщики, 3 крейсера и 6 миноносцев. С 9 сентября начались бои за остров Муху. Утром 14 сентября вражеские войска одновременно высадились на северной и южной оконечностях Муху. Гарнизон острова оказал противнику упорное сопротивление, однако врагу удалось закрепился. Трое суток на острове продолжались тяжелые, непрерывные бои. Оборонявшиеся несли большие потери, и 17 сентября они отошли на Сааремаа.
Началась не менее ожесточенная борьба на острове Сааремаа. Изнурительные, непрерывные бои с превосходящими силами противника вынуждали наши подразделения постепенно отходить к полуострову Сырве. Этот рубеж держали до 30 сентября. Враг бросал в атаки свежие части пехоты при сильной поддержке авиации и даже легких крейсеров «Лейпциг», «Эмден» и миноносцев. Точные залпы наших береговых батарей под командованием капитана А.М. Стебеля (батарея № 315 мыс Церель) и военкома батареи № 43 старшего политрука Г.А. Карпенко (мыс Рахусте), а также атаки торпедных катеров заставили немецкие корабли уйти.
Атака моряков
C плаката С.С. Боим
К этому времени в рядах защитников полуострова осталось всего около полутора тысяч человек, к концу подходили боеприпасы. Поэтому было принято решение оставить полуостров Сырве, а его защитников перебросить на остров Хийумаа. Однако из-за штормовой погоды и сильного противодействия врага эвакуировать удалось далеко не всех. 4 октября в Москве была принята последняя радиограмма с Сааремаа: «Иду в бой, в последний бой. Прощайте, прощайте…» В 16 часов 10 минут связь с героическими защитниками Сааремаа прервалась.
Немногочисленный гарнизон острова Хийумаа под командованием полковника А.С. Константинова усиленно готовился к обороне, но сил и средств у них было очень мало. Береговая батарея № 44 под командованием старшего лейтенанта М.А. Катаева целый день вела бой в полном окружении. С наступлением темноты артиллеристы гранатами расчистили себе путь на север. Раненый сержант Е.Ф. Попов добровольно остался на батарее и взорвал ее вместе с ворвавшимися гитлеровцами.
Не меньшее мужество и стойкость защитники острова проявили и на других позициях. Только на четвертые сутки они отошли к мысу Тахкуна. Здесь было получено приказание эвакуировать личный состав на Ханко. До 22 октября удалось эвакуировать 570 человек. Оставшаяся советские воины бились до конца. Об этом красноречиво говорит и подвиг матроса Николая Чижа, до последней возможности защищавшего даже верхнюю площадку маяка Тахкуна. Беспримерное мужество и стойкость в обороне являются одной из важнейших боевых традиций нашего флота. Наглядным примером этого служит оборона острова Тенедос в 1807 году. Подробности читайте здесь. И все-таки, более 15 тысяч наших бойцов и командиров, израненных, контуженных, смертельно измученных непрерывными боями за Моонзунд, попало в плен.
Свыше 40 суток продолжалась оборона Моонзундских островов. Их защитники прекрасно понимали, что находятся в глубоком тылу врага, за сотни километров от линии фронта и отступать им некуда. Но они мужественно сражались твердо зная, что каждый убитый ими фашист уже никогда не пойдет в атаку под Ленинградом или Москвой, не будет штурмовать Одессу или Мурманск. Защитники Моонзунда отвлекли на себя свыше 50 тысяч фашистов, половина из них навсегда осталась в каменистой земле архипелага. В ходе боев было уничтожено более 20 судов, 41 самолет и много другой техники.
Судьба командира
Трагически сложилась и судьба их командира – генерал-лейтенанта береговой службы Алексея Борисовича Елисеева. Он начинал свою службу на флоте в 1908 году и был хорошо известен как балтийский матрос-большевик, активный участник Гражданской войны, большой специалист в области береговой артиллерии. В апреле 1938 года комдив Елисеев был арестован и под пытками «признал» участие в «военно-фашистском заговоре». К счастью, суд оправдал его, и после 18 месяцев заключения, издевательств и пыток Алексей Борисович вышел на волю и был восстановлен на флоте.
Вскоре после завершения боев на Моонзунде Елисеев прибыл в Ленинград и был назначен на малозначащую должность. Он воспринял это очень болезненно, полагая, что его знания и боевой опыт, могли бы принести больше пользы. Наконец, его вызвали в Москву. Этот вызов он воспринял с тяжелым сердцем, поскольку не исключал возможность ареста. В то время уже сидели в застенках бывший командующий Морской обороной Ленинграда контр-адмирал К.И. Самойлов и адмирал Г.И. Левченко, которого сухопутные начальники сделали «козлом отпущения» за поражение в Крыму. Не менее показательна в этой связи и трагическая судьба адмирала Галлера.
Он хорошо помнил об издевательствах НКВД, и не хотел их повторения. Тяжело было видеть и то, что защитников Ханко в Ленинграде встречали как героев, а о моонзундцах вообще забыли. Переживания за судьбу оставленных в плену подчиненных и моральное воздействие завистников, которые обвиняли его в преждевременной сдаче противнику столь важных для обороны островов, сломили волю Алексея Борисовича. Он не стал ждать нового ареста и не стал отвечать на обвинения, 21 декабря 1942 года генерал-лейтенант А.Б. Елисеев застрелился.
В наше время добрая память о подвиге героев Моонзунда и об их командире Алексее Борисовиче Елисееве восстановлена.
При написании статьи были использованы следующие материалы:
- Чернов Ю.И. Война погасила маяки. Москва. 1985 г.
- Кузнецов Н.Г. Курсом к победе. Москва. 2003 г.
- Чернышев А.А. Балтийский флот в битве за Ленинград. Москва. 2014 г.
- Близниченко С.С. К 125-летию со дня рождения генерал-лейтенанта береговой службы А.Б. Елисеева. Журнал «Военно-исторический архив» № 3 за 2012 г.
Каменистая земля Моонзунда обильно полита кровью наших воинов, мужественно сражавшихся здесь на протяжении многих веков. Их подвиги бережно хранятся в памяти народной, но они еще очень слабо отражены в нашей литературе. Эта статья всего лишь скромное напоминание о них. Вы, уважаемый читатель, можете дополнить этот рассказ яркими примерами или своей оценкой произошедшего. Напишите об этом в комментариях. Это будет интересно всем!
Эстония. Таллинн. Мыс Церель — ВолгаПитер
Начитавшись и насмотревшись Пикуля, решили посетить места боевой славы Русского Флота. А именно, мыс Церель, посмотреть на Ирбенский пролив,….
Итак, в путешествии приняли участие: командир экипажа – Кирилл (сын), штурман – Надежда (жена), пилот – я (папа и муж).
Выезжали в субботу, рано утром (хотели в семь, получилось в восемь). Дождь, провожал нас до самой границы. В Ивангороде при торможении заглох, потом еще раз на заправке. Подложил гаечку. Поехали.
Границу прошли более или менее нормально. Часа два примерно потратили на стояние в очередях. Надо выезжать раньше. Вроде должно быть оптимально часиков в шесть в субботу. Тогда есть вероятность проскочить между двумя волнами – теми кто уезжает в пятницу и теми, кто уезжает в субботу. Но есть одно но – торговый комплекс «Астри», где рекомендую позавтракать, открывается в 10 (9 по Москве).
Эстонцу не понравилось мое подспущенное переднее колесо, но заворачивать нас не стал, за что спасибо.
Сын, при проверке документов и печати на выезд на весь пункт спросил у пограничницы: «А в зоопак мозя? Там соон! И лев стасий, вот с такими зубами!» (А в зоопарк можно? Там слон! И лев страшный, вот с такими зубами!).
Как только пересекли границу дождь утих, выглянуло солнышко и стало припекать. После плотного и недорого завтрака в «Астри» пошли в сторону Таллинна.
Теперь о коварстве эстонских дорог. Они хорошие, а при ограничении 90 просто отличные. А тут еще солнышко прямо на черную приборную панель, ровный шум мотора. И никого. Короче, я задремал и приехал правым бортом в ограждение – царапина, но не приятно. Две кружки кофе и адреналин позволили дальше добраться без приключений. Это я к чему? К тому, что НАДО останавливаться на заправке «Олерекс», на развилке Таллин — Нарва Йыесу (игрек образный перекресток, заправка слева).
Итак, трасса Нарва – Таллинн. Очень прошу не гнать, не потому что чревато штрафами, а просто что бы была возможность глядеть не только на дорогу, но и по сторонам.
И увидеть танец аистов. Вообще аистов в Эстонии много, машин они не боятся, так что прошу смотреть и за крылатыми пешеходами.
Таллинн.
Город наполненный запахом корицы. Дышите в старом городе глубже. История, история… Таллинн может считаться полным отображением поговорки «было бы счастье, да несчастье помогло». Эстония всегда была самой бедной из Прибалтийский государств, поэтому когда в 17-18 веках старинные здания стали считаться немодными и уродующими облик города, государства выкупало эти дома и сносило их по самый фундамент. А у Эстонии просто не было на это средств…. Поэтому старинных домов в Таллинне так много. По поводу вождения в городе – очень много улочек с односторонним движением и сами улочки очень узкие. Припарковаться где-нибудь в бесплатном месте очень сложно. Но есть выход – в R-киоске покупаем парковочные часы и ищем стоянку со стилизованным изображением часов и надписью времени (30 min, 3 h). Указанное время означает сколько можно стоять бесплатно, выставляем время на парковочных часах и стоим. По окончании времени переезжаем на другое место и опять выставляем часы. В Старом Городе все парковки платные.
Что посетить? Поскольку в Эстонии запрещено курить в общественных местах, то рекомендации одинаковы для детей и взрослых.
Итак, первый эстонский паб «Хелл Хант» около ратушной площади, также недалеко от площади около маленького красного домика, рядом с чесночным рестораном «Бальтазар» есть что-то вроде кафе (условно назовем «стекляшка»). Кроме этого, «Олде Ханса», «Пепперсак» — знаменитые аутентичные рестораны Таллинна.
Но……
Но ребенок тянул в зоопарк. Если Вы в Таллинне надолго, то приобретите в туристическом центре «Таллинн Карт» и зоопарк будет бесплатно, как и многие музеи. Зоопарк большой, так что если ребенок маленький, то рассчитывайте на два дня.
Узнаете? Как нет?? Это же Беркут собственной персоной!
Но горн звал вперед.
До паромной переправы Виртсу добрались без приключений. Правда пришлось проползти по щебенке (дорогу ремонтировали). Вы когда-нибудь видели выпученные глаза водителя «Мустанга»? Вооооот, а я видел, его пузотерка шла навстречу, водителю было реально страшно…… по щебню-то, где каждый камень так и грозит поцарапать…
Не узнаете? Это же торфяные болота. Да, те самые…. Ночью погулять не хотите? А то могу устроить…. Собачку-то, говорят, забыли после съемок.
Эстонцы специально срубили ветрозащитную полосу на участке дороги, чтобы туристы могли своими глазами увидеть Гринменскую трясину.
Итак, мы переправились на остров Муху (бывший Даго) через пролив Моон, и фарватер «Славы». «Слава» русский линкор затопленный из-за повреждений полученных в бою в германским флотом в Рижском заливе закрывший путь немцам к Петрограду.
А это наша следующая остановка. Осмусаарская дамба. Единственный путь отступления русской армии с острова Эзель. Оборонял дамбу «Батальон Смерти» из моряков Балтийского флота. Покажите эту фотографию пожилому немцу и посмотрите как он бледнеет от ужаса.
Но дальше..
Ирбены. Пожалейте немца и не показывайте ему эту фотографию. Вообще эстонцы (сааремцы) что в Первую, что во Вторую оказались между молотом и наковальней поэтому:
Памятный знак на командном пункте батарей Стебеля – это уже Великая Отечественная. До самой батарей не добрались. Весь остров заболоченный, так что дальше чем триста метров в лес забираться я не рискнул.
А это уже местечко Охисааре – там очень много плоских камней и влюбленные со всей Эстонии приезжают сюда для того чтобы сложить такую пирамиду на счастье.
Озеро Медвежье – старейшее озеро Эстонии.
Итак, после этого мы взяли курс домой.
По поводу отношения эстонцев к русским. В городе Куресааре я окончательно залил ЭБУ и машина встала полностью. После получаса стояния с открытым капотом ко мне подошел мужчина и спросил «Сломался?» на что мне оставалось только кивнуть.Никогда не угадаете кто это был! Волговод! В армии возил командира на 24. Короче он мне вызвал сервис, отвез жену с ребенком в гостиницу, меня в магазин за продуктами…… Был одарен календариком (от денег отказался). В сервисе (хозяин тоже волговод) высушили мозги, заменили прокладку на выхлопе. Всего 84 евро. Я не знаю чего они сделали, но на полубаке я проехал около 400 километров.
Если кого-нибудь интересует что-нибудь еще, то с радостью расскажу более подробно.
Единственный минус — 5 часов ожидания выезда в Россию. А плюс — эстонские полицейские взявшие под козырек.
Фотографии в альбоме. Почему-то не вставляются
ФОТО читателя Delfi: Тайны моонзундских батарей
Рывок на юг острова. В Аренсбурге, т.е. Курессааре подхватываем моих сааремаских друзей Свету и Андрея. Андрей, немного зная мой неугомонный нрав, не удивляется тому, что мы едем смотреть батареи на самую южную оконечность острова. Потому что российская история, потому что Пикуль, потому что…
Проскочили Мяндьяла и движемся уже по полуострову Сырве. На пригорке два памятника — советским солдатам, погибшим во Второй мировой войне, и немецким солдатам, погибшим в той же войне. Земля приняла всех убитых.
Вот и остатки 43-й (Церельской батареи). Начало постройки батареи — осень 1916 года, а в строй батарея вступила в апреле 1917 года.
“Церель открыл огонь, после чего германские дредноуты начали перемешивать его с землей. Высоко взлетели разбитые рельсы. Дым вздымался на чудовищную высоту. Осколки жестоко изрезывали бетон укрытий. Из каркасов брустверов уродливо выпучивало основу досок и бревен… Церель сражался”.
В.Пикуль, “Моонзунд”
Командовал батареей Николай Сергеевич Бартенев, который стал прообразом главного героя романа Валентина Пикуля “Моонзунд” — старшего лейтенанта Артеньева. А строил батарею выдающийся фортификатор Н.И.Унгерман.
“Близкий разрыв засыпал его мерзлой землей, сверху упала мертвая чайка. Шатаясь, он снова приник к прицелам, и перед старшим лейтенантом — какой уже год! — все так же волновалась серая простыня Ирбен, на которой ползали, будто вши, отвратительные живчики вражеских кораблей. Время от времени оттуда вспыхивали огоньки, почти мирные, похожие на булавочные головки, — это были выстрелы дредноутов, которые отзывались на Цереле страданием…”
Закат Церельской батареи произошел в 1917 году. Орудия батареи не имели кругового сектора обстрела и не могли противодействовать войскам, наступавшим на мыс Церель по полуострову Сворбе. Одновременно позиции батареи подвергались бомбардировкам с воздуха. После очередной воздушной атаки 1 октября 1917 года, загорелся склад пороховых зарядов, в результате последующего взрыва погиб 121 человек. 43-я батарея держалась до 15 (2) октября. Под огнем немецких линкоров орудия и погреба были взорваны, и личный состав покинул батарею.
Мы отправляемся к маяку. Где-то за спиной остается Сааремаа, да и вся Эстония. До Латвии рукой подать. Вот она, на горизонте. Мобильные телефоны тут же дружно переходят на латвийский роуминг. Странно, ты вроде бы в Эстонии, а говорить приходиться через латвийскую мобильную сеть. Узким кончиком языка вклинивается полуостров Сырве в Ирбенский пролив.
В 1646 году генерал-губернатор Лифляндии Габриель Оксенштерн велел построить здесь примитивный маяк — светящийся знак. В 1770 году на южной оконечности полуострова Сырве была построена каменная башня, разрушенная во время первой мировой войны. шняя коническая монолитная железобетонная башня построена в 1960 г. Высота Маяка Сырве от уровня земли составляет 52 метра, от уровня моря 53 метра.
Июнь. Скоротечно северное лето. А уж погожих дней вообще по пальцам перечесть. Навстречу солнцу раскрылись все растения острова, наполняя ароматом морской воздух. Эскадрильи пчел без устали атакуют медоносные аэродромы цветов.
Тишина и умиротворенность. Какое-то безвременье.
На обратном пути пытаемся проехать еще к одной батарее, 315-й батарее под командованием Стебеля Александра Моисеевича. Главной задачей батареи — не допустить прорыва кораблей противника в Рижский залив, через Ирбенский пролив. Работы по строительству начались в 1940 году. В 1941 году батарея была взорвана. Следует заметить, что командный пункт 315-й батареи был расположен точно на позиции батареи №40 времен Первой мировой войны.
В лес ведет дорога, по которой машины точно не проедут. Идем вперед пешком. Места становятся труднопроходимыми. Андрей замечает cвежие следы крупного кабана. Дальше идти небезопасно.
Моонзунд хранит свои тайны. И хранит свою историю.
“Долг, честь, присяга — это ведь не пустые слова!
Нельзя их закидывать под лавку…”
В.Пикуль, “Моонзунд”
Сигнальные рецепторы лектинов C-типа, распознавание микробов и иммунитет
Резюме
Сигнальные рецепторы лектинов C-типа (CLR) имеют решающее значение в формировании иммунного ответа на грибковые патогены, но сравнительно мало известно о роли этих рецепторов в бактериальной среде. , вирусные и паразитарные инфекции. CLR выполняют множество разнообразных функций в зависимости от сигнальных мотивов в их цитоплазматических доменах и могут вызывать эндоцитарные, фагоцитарные, антимикробные, провоспалительные или противовоспалительные реакции, которые являются либо защитными, либо нет во время инфекции.Понимание роли CLR в формировании противомикробного иммунитета предлагает большой потенциал для будущей разработки терапевтических средств для лечения заболеваний. В этом обзоре мы сосредоточимся на распознавании бактериальных, вирусных и паразитарных патогенов CLR и на том, как эти рецепторы влияют на исход инфекции. Мы также кратко расскажем о роли CLR в противогрибковом иммунитете.
Введение
Врожденная иммунная система обеспечивает первую линию защиты от микробной атаки и индуцируется распознаванием микробных компонентов, известных как патоген-ассоциированные молекулярные паттерны (PAMP) или связанные с микробами молекулярные паттерны (MAMP), по паттерну. рецепторы распознавания (PRR).PAMP высоко консервативны и, как правило, уникальны для микробов (Akira et al ., 2006). Грибковые PAMPs состоят в основном из углеводных структур клеточной стенки, тогда как бактериальные PAMPS варьируются от липопротеинов, липополисахаридов (LPS), флагеллина и пептидогликана до структур бактериальных нуклеиновых кислот. С другой стороны, вирусы в основном распознаются по их нуклеиновым кислотам, таким как двухцепочечная (дцРНК) или одноцепочечная РНК (оцРНК) и вирусная ДНК, хотя также могут распознаваться гликопротеины поверхностной оболочки.Хотя PAMP не так хорошо охарактеризованы, паразиты, особенно гельминты, такие как Schistosoma mansoni и Trichuris muris , или простейшие, такие как Leishmania infantum и Plasmodium berghei , также распознаются PRR млекопитающих (McGetuinness 9000). al ., 2003; Broz, Monack, 2013; Drummond, Brown, 2013).
PRR распознавание PAMP может привести к активации внутриклеточных сигнальных путей, которые вызывают врожденные ответы против патогенов и направляют развитие адаптивного иммунитета.Также важно упомянуть распознавание связанных с повреждениями молекулярных паттернов (DAMP) с помощью PRR. Молекулы, высвобождаемые стрессовыми клетками или клетками, подвергающимися некрозу, могут действовать как сигналы опасности и способствовать воспалительным ответам (см., Например, Yamasaki et al ., 2008). Важными здесь PRR являются сигнальные трансмембранные лектиновые рецепторы С-типа (CLR), которые, как широко признано, играют важную роль в противогрибковом иммунитете (см. Hardison and Brown, 2012 для недавнего обзора).Менее признана их роль в иммунитете к другим микроорганизмам. Поэтому этот обзор будет сосредоточен на роли сигнальных CLR в иммунитете к бактериям, вирусам, гельминтам и простейшим и лишь кратко обсудим самые последние достижения в нашем понимании их роли в противогрибковом иммунитете. Кроме того, мы включили некоторые важные CLR, в которых сигнальный путь неясен, особенно из группы VI. Структуры выбранных CLR, которые будут обсуждаться, представлены на рис..
Рецепторы лектинов C-типа: общий обзор
CLR включают большое семейство рецепторов, которые связывают углеводы через один или несколько доменов распознавания углеводов (CRD) или которые обладают структурно схожими лектин-подобными доменами C-типа (CTLD), которые не имеют обязательно узнайте углеводные лиганды. CLR делятся на 17 групп в зависимости от особенностей, включая филогению и структуру (Зеленский и Греди, 2005). В этом обзоре обсуждаются CLR из группы II (кальций-зависимые лектины с одним CRD), группы V (кальций-независимые рецепторы с одним CTLD) и группы VI (кальций-зависимые лектины с несколькими CRD) (см. Таблицу).Основываясь на их сигнальном потенциале, CLR могут быть далее подразделены на (i) активирующие Syk-связанные CLR с доменами иммунорецепторного тирозинового мотива активации (ITAM), (ii) ингибирующие CLR с доменами иммунорецепторного тирозинового мотива ингибирования (ITIM) или ( iii) CLR без четких доменов ITAM или ITIM, таких как MR, DEC ‐ 205 и DC ‐ SIGN (Zelensky and Gready, 2005; Sancho and Reis e Sousa, 2012).
Таблица 1
Избранные CLR, упомянутые в этом обзоре
| CLR | Лиганды | Происхождение лиганда | Избранные ссылки | |
|---|---|---|---|---|
| Группа II: Зависимый от кальция CRD | Dectin ‐ 2 | α-маннаны O-связанный маннобиоз-богатый гликопротеин | М.туберкулез S. mansoni SEA C. albicans Malassezia видов. Аллергены HDM | Риттер и др. ., 2010; Мивиссен и др. ., 2012; Исикава и др. , 2013; Prasanphanich et al ., 2013; Салазар и др. ., 2013 г. |
| CLECSF8 | TDM | M. bovis К.пневмония | Miyake et al ., 2013; Steichen et al ., 2013 | |
| Mincle | α-манноза маннит-связанный глицерогликолипид маннозил жирных кислот TDM | M. tuberculosis C. albicans Malassezia видов. | Ishikawa et al ., 2009; Ли и др. ., 2012; Санчо и Рейс и Соуза, 2012 г .; Исикава и др. ., 2013 г. | |
| DC ‐ SIGN | Высокое содержание маннозы SlpA | ВИЧ ‐ 1 Корь Денге Mycobacterium spp. Грипп A SEA Leishmania spp. Хеликобактер пилори Lactobacillus spp. | Gringhuis et al ., 2007; Константинов и др. ., 2008; Гейтенбек и Гринхейс, 2009 г .; Месман и др. ., 2012; Avota и др. ., 2013; Чен и др. ., 2013; Харман и др. ., 2013; Hillaire et al ., 2013 | |
| SIGNR3 | Высокоманноза и фукоза | L. infantum SEA | Powlesland et al ., 2006; Мивиссен и др. ., 2012; Лефевр и др. ., 2013; Prasanphanich et al ., 2013 | |
| SIGNR1 | Высокоманноза и фукоза | SEA | Galustian et al ., 2004; Мивиссен и др. ., 2012; Prasanphanich et al ., 2013 | |
| MGL | Lewis X | SEA | Van Vliet et al ., 2005; Мивиссен и др. ., 2012; Tundup и др. ., 2012 г. | |
| DCIR | неизвестно | HIV ‐ 1 | Sancho and Reis e Sousa, 2012 | |
| Группа V: кальцийнезависимый не-CRD | Dectin-1 | β-глюканы | Л.младенец C. albicans Mycobacterium spp. | Хардисон и Браун, 2012 г .; Лефевр и др. ., 2013 |
| CLEC5A | Неизвестно | Вирус денге JEV | Chen et al ., 2012; Драммонд и др. ., 2013 | |
| DNGR ‐ 1 (CLEC9A) | F ‐ actin | Вирус осповакцины Вирус простого герпеса | Иборра и др. ., 2012; Зеленай и др. ., 2012 | |
| Группа VI: кальций-зависимые множественные CRD | Маннозный рецептор (MR) | Высокое содержание маннозы Омега ‐ 1 МанЛам | SEA Trichuris muris Мезоцестоидес корти Аллергены HDM Mycobacterium spp. К. пневмония S. pneumonia F. tularesis | Kang et al ., 2005; Deschoolmeester et al ., 2009; Гейтенбек и Гринхейс, 2009 г .; Everts et al ., 2012; Мивиссен и др. ., 2012; Мишра и др. ., 2013; Prasanphanich et al ., 2013; Салазар и др. ., 2013 г. |
| DEC ‐ 205 (CD205) | PLA | Ю.pestis | Zhang et al ., 2008 г. |
Эта статья размещена в свободном доступе через PubMed Central в рамках программы реагирования на чрезвычайную ситуацию в области общественного здравоохранения COVID-19. Его можно использовать для неограниченного повторного использования в исследованиях и анализа в любой форме и любыми средствами с указанием первоисточника на время чрезвычайной ситуации в области общественного здравоохранения.
Активация CLR может индуцировать внутриклеточные сигнальные пути двумя способами: во-первых, через непрямую передачу сигналов, когда рецепторы, такие как индуцируемый макрофагами лектин C-типа (Mincle или CLEC4E), Dectin-2 (CLEC6A) и семейство лектиновых доменов C-типа 5A (CLEC5A) связывается с ITAM, содержащим адаптерные молекулы, такие как γ-цепь рецептора Fc (FcRγ) или DAP12.
Второй механизм, используемый Dectin-1 (или CLEC7A) и DNGR-1 (CLEC9A), включает прямую передачу сигналов через ITAM-подобные мотивы, расположенные в цитоплазматическом хвосте этих рецепторов (Rogers et al ., 2005; Geijtenbeek and Gringhuis , 2009). Передача сигналов через оба механизма включает привлечение к фосфорилированным тирозиновым остаткам тирозинкиназы селезенки (Syk), которая, в свою очередь, координирует комплекс CARD9, B-клеточной лимфомы 10 (Bcl10) и белка транслокации 1 лимфоидной ткани, ассоциированной со слизистой оболочкой (Malt1). .Более того, ассоциированный с апоптозом спек-подобный белок, содержащий CARD (ASC), также, как было показано, фосфорилируется Syk- и Jnk-зависимым образом (Hara et al ., 2013). Протеинкиназа C-δ (PKCδ) также является важным элементом в этом пути (Strasser et al ., 2012). Передача сигналов от Dectin-1 и др. Лектинов также включает дополнительные пути, такие как Syk-независимая активация Raf-1 (Gringhuis et al ., 2009). Эти сигнальные пути приводят к активации нескольких нижестоящих молекул, включая NFκβ и митоген-активируемые протеинкиназы (MAPK), в конечном итоге вызывая клеточные ответы, включая фагоцитоз, созревание ДК, хемотаксис, респираторный взрыв, активацию инфламмасом и продукцию цитокинов и других медиаторов ( Драммонд и др. ., 2011; Штрассер и др. ., 2012). Более того, эти рецепторы также могут управлять развитием адаптивного иммунитета, в частности Т-хелпер (Th) 1 и Th27 CD4 + и CD8 + адаптивные ответы Т-клеток, хотя некоторые CLR также способны управлять Th3-иммунитетом (Kerrigan and Brown , 2011; Sancho, Reis e Sousa, 2012).
В то время как передача сигналов через ITAM-несущие рецепторы приводит к активации клеток, ITIM-несущие рецепторы обычно опосредуют тормозные функции. Например, DCIR содержит ITIM в своем цитоплазматическом хвосте, который привлекает тирозинфосфатазы (SHP-1 и SHP-2) после связывания лиганда для модуляции сигнальных путей, индуцированных другими PRR, таких как ингибирование индуцированной TLR8- или TLR9 продукции цитокинов ( Geijtenbeek and Gringhuis, 2009).Однако функция ингибирующих CLR не всегда четко определена, поскольку эти рецепторы также могут выполнять функции активации, тем самым опосредуя активацию клеток. Они могут либо рекрутировать новые субстраты в свои цитоплазматические домены, либо ингибировать другие ингибирующие рецепторы (Redelinghuys and Brown, 2011).
Роль лектинов C-типа в распознавании микробов и иммунитете
Бактерии
Роль CLR в антибактериальном иммунитете лучше всего охарактеризована для M. tuberculosis (MTB).Множественные CLR, включая Dectin-1, DC-SIGN, рецептор маннозы (MR) и Mincle, участвовали в контроле MTB in vitro , однако каждый из этих CLR, по-видимому, является избыточным в контроле инфекции in vivo . Например, было показано, что Dectin-1 необходим для продукции IL-12p40 селезеночными ДК в ответ на микобактерии in vitro , но был избыточным во время заражения in vivo (Rothfuchs et al ., 2007; Court и др. ., 2010; Маракалала и др. ., 2010). MR связывается с липоарабиноманнаном, ограниченным маннозой (ManLAM) на поверхности M. tuberculosis , опосредуя бактериальный фагоцитоз и ограничивая слияние фагосома-лизосома в макрофагах, но потеря MR не повлияла на антимикобактериальный иммунитет in vivo09 (et Schlesinger al ., 1994; Kang et al ., 2005; Court et al ., 2010). Mincle опосредует распознавание микобактериального пуповинного фактора, трегалоза-6,6′-димиколата (TDM), и было показано, что он играет важную роль в управлении иммунными ответами на TDM in vivo , включая образование гранулем (Ishikawa et al ., 2009). Тем не менее, мыши с нокаутом Mincle имели нормальные гранулемы и не демонстрировали явных дефектов во время инфицирования живым MTB (Lee et al ., 2012; Heitmann et al ., 2013). DC-SIGN распознает ManLAM и α-глюкан, предотвращая созревание DC и производство IL-10 (Geijtenbeek et al ., 2003; Geurtsen et al ., 2009). Хотя у мышей с дефицитом мышиного гомолога DC-SIGN, SIGNR3 — / — были обнаружены дефекты в раннем контроле MTB, у них был выявлен эффективный анти-микобактериальный адаптивный иммунный ответ с гранулематозными поражениями, сопоставимыми с контрольными животными дикого типа (Tanne et al. ., 2009).
Несмотря на очевидную избыточность этих рецепторов in vivo , общий нисходящий сигнальный путь CLR с участием CARD9 имеет решающее значение для защиты, поскольку мыши CARD9 — / — присутствуют с неконтролируемой бактериальной репликацией и обостряются нейтрофильным воспалением легких, за которым следует смерть (Dorhoi et al ., 2010). Это говорит о том, что комбинация CLR или неизвестная CLR важна для защиты от MTB. Недавние исследования показали, что Mincle может образовывать рецепторный комплекс с CLECSF8 (CLEC4D или MCL) и FcεRIγ, и предполагается, что этот гетеротримерный комплекс является функционально оптимальной формой для этих CLR (Lobato-Pascual et al ., 2013). Однако роль CLECSF8 в заражении живым MTB не определена.
Недавно было показано, что CLECSF8 играет важную роль в разрешении пневмонии, вызванной Klebsiella pneumoniae (Steichen et al ., 2013). Мыши CLECSF8 — / — были более восприимчивы к легочному сепсису, чем мыши дикого типа, с повышенной бактериальной массой, гипервоспалением и тяжелой патологией легких, которые коррелировали с массивным накоплением нейтрофилов. Эти результаты предполагают, что CLECSF8 играет важную роль в разрешении воспаления, и является первым отчетом, описывающим физиологическую функцию этого CLR.
Помимо M. tuberculosis , DC-SIGN также взаимодействует с широким спектром других бактериальных патогенов, включая M. leprae , Helicobacter pylori и виды Lactobacillus (Geijtenbeek and Gringhuis, 2009). Было показано, что Lactobacillus reuteri и L. casei связывают DC-SIGN и индуцируют регуляторные Т-клетки (Smits et al ., 2005), в то время как белок поверхностного (S) слоя (SlpA) на поверхности L. acidophilus был идентифицирован как лиганд этого CLR (Константинов и др. ., 2008). MR также распознает ряд других видов бактерий, экспрессирующих маннозу, включая M. kansasii , K. pneumonia , Streptococcus pneumoniae и Francisella tularensis . Однако MR, по-видимому, не важен во время инфицирования этими патогенами in vivo (Geijtenbeek and Gringhuis, 2009).
Было также показано, что другие, менее хорошо охарактеризованные CLR распознают бактерии. DEC-205, например, является членом семейства рецепторов маннозы и связывается с активатором плазминогена (PLA) на поверхности Yersinia pestis , который опосредует прикрепление бактерий (Lähteenmäki et al ., 1998). Было обнаружено, что этот CLR способствует распространению этого патогена, и это имело пагубные последствия для хозяина (Zhang et al ., 2008).
Вирусы
Вирусы — это многочисленные, быстро эволюционирующие патогены, которые создают постоянную угрозу для иммунной системы хозяина. В отличие от преимущественно защитных реакций, которые CLR опосредуют на другие патогены, распознавание вирусов CLR имеет тенденцию способствовать передаче, инфекции и воспалению. Распознавание ВИЧ с помощью DC-SIGN является хорошо известным примером пагубного эффекта передачи сигналов CLR в ответ на вирус.Во-первых, связывание ВИЧ-1 с DC-SIGN не только модулирует TLR-индуцированную продукцию IL-10 посредством передачи сигналов через Raf-1, но также нарушает пролиферацию Т-клеток и TLR-индуцированное образование дендритов DC (Gringhuis et al ., 2007). Во-вторых, gp120 ВИЧ облегчает DC-SIGN-опосредованное проникновение вируса в клетки, приводит к инфицированию CD4 + клеток-мишеней (Harman et al ., 2013) и ускоряет апоптоз DC, что отрицательно влияет на созревание DC, что обычно способствует распознаванию патогенов. иммунной системой (Chen et al ., 2013). Следовательно, предотвращение связывания ВИЧ с DC слизистой оболочки путем блокирования CLR может потенциально предотвратить передачу ВИЧ.
Лиганды DC-SIGN не ограничиваются ВИЧ, но включают ряд других вирусов, таких как цитомегаловирус, вирус денге, вирус Эбола, вирус гепатита С, коронавирус SARS-коронавирус, вирус Западного Нила и вирус кори (Mesman et al ., 2012; Avota et al ., 2013; Hillaire et al ., 2013). Совсем недавно было показано, что DC-SIGN способствует репликации вируса гриппа A путем связывания с гликанами на гемагглютинине, способствуя связыванию вируса с клетками и интернализации in vitro (Hillaire et al ., 2013). Однако вклад DC-SIGN зависел от степени гликозилирования вирусного гемагглютинина.
Вирусное использование CLR также может индуцировать провоспалительные цитокины, ведущие к тяжелой патологии для хозяина. Ранее было показано, что MR и CLEC5A связывают вирус денге in vitro , что приводит к инфицированию макрофагов и активации инфламмасом соответственно. В отличие от обычных CLR, которые участвуют во проникновении вируса денге в клетки-мишени, CLEC5A регулирует индуцированные вирусом провоспалительные цитокины и блокирование передачи сигналов, опосредованной CLEC5A, ослабляет продукцию провоспалительных цитокинов инфицированными макрофагами, снижая смертность и поддерживая иммунитет хозяина, что приводит к разрешению инфекции.Это указывает на то, что CLEC5A имеет решающее значение в регулировании воспалительных реакций, вызванных патогенами. Повышенные уровни TNF-α во время инфекции были связаны с активацией DAP12, предполагая, что CLEC5A напрямую взаимодействует с вирионом денге (Chen et al ., 2008). Точно так же вирус японского энцефалита (JEV) также напрямую связывает CLEC5A и вызывает нейровоспаление через активацию DAP12 в макрофагах. Блокирование CLEC5A снижает повреждение нейронов, секрецию провоспалительных цитокинов, проницаемость гематоэнцефалического барьера и клеточную инфильтрацию в центральную нервную систему (ЦНС) in vivo (Chen et al ., 2012). Вместе эти исследования предполагают, что блокада CLR может облегчить повреждение тканей и увеличить выживаемость пациентов с воспалительными заболеваниями, вызванными вирусами.
В отличие от этих пагубных ролей, CLR также могут вызывать защитные реакции. Примером этого является DNGR-1 (CLEC9A) в борьбе с вирусами осповакцины (Iborra et al ., 2012) и вирусом простого герпеса (Zelenay et al ., 2012). DNGR-1 экспрессируется подмножеством DC и обнаруживает мертвые клетки, способствуя перекрестной презентации антигена Т-клеткам CD8 + .Хотя DNGR-1-дефицитные DC активируются после взаимодействия с инфицированными вирусом клетками, они больше не способны к перекрестной презентации антигенов. Это приводит к более слабым ответам Т-клеток CD8 + , замедленному разрешению поражения и более высокой вирусной нагрузке, что позволяет предположить, что восприятие повреждения ткани с помощью DNGR-1 является ключевым компонентом противовирусного иммунитета.
Другим примером защитных ответов, опосредованных CLR, является роль DCIR во время заражения вирусом Чикунгунья (Long et al ., 2013). У мышей DCIR — / — развилось более тяжелое воспалительное заболевание с асимметричным цитокиновым ответом как in vivo , так и in vitro . Таким образом, ингибирующие функции этого рецептора играют важную роль в подавлении патологических воспалительных реакций, вызванных этим патогеном (Long et al ., 2013).
Гельминты
Гельминты-паразиты направляют CD4 + Th-клетки хозяина на Th3 и противовоспалительные реакции, а также вызывают альтернативную активацию макрофагов.Считается, что гликаны гельминтов играют решающую роль в управлении этими реакциями, учитывая большое количество гликановых фрагментов, которыми они обладают; вполне вероятно, что многие CLR участвуют в иммунитете к этим патогенам. Растворимый яичный антиген (SEA) Schistosoma mansoni cercariae был первым описанным паразитоспецифическим лигандом для DC-SIGN (Meyer et al ., 2005). Впоследствии гликопротеины из SEA нескольких видов шистосом ( S. mansoni , S. hematobium , S.japonicum ) были описаны как лиганды для DC-SIGN (van Die et al ., 2003).
Другие сигнальные CLR, как было показано, распознают SEA или гликанов S. mansoni , хотя не все фактические лиганды были идентифицированы. Среди них Dectin-2 распознает неизвестный лиганд в SEA; SIGNR1, SIGNR3 и MR распознают Lewis x и высокоманнозные N-гликаны; и макрофагальный галактозный лектин (MGL) распознает Lewis x, LDN и LDN-F в SEA (Meevissen et al ., 2012; Prasanphanich et al ., 2013). Более того, MR распознает Omega-1 в SEA и обусловливает DC для прайминга Th3 (Everts et al ., 2012). Dectin-2, в частности, был первым Syk-связанным CLR, который был связан с гельминтными инфекциями, и играет роль в регуляции иммунных ответов гельминтов путем косвенного снижения Th3-опосредованной патологии. Здесь было обнаружено, что Dectin-2 индуцирует активную секрецию IL-1β путем активации инфламмасомы Nlrp3 в ответ на S. mansoni SEA (Ritter et al ., 2010).
Другие CLR также участвуют в противогельминтном иммунитете, но их роль in vivo менее ясна. Например, мышиный SIGNR1 (гомолог DC-SIGN) связывает SEA in vitro , но мыши SIGNR1 — / — дают нормальный ответ во время инфицирования S. mansoni . И MGL, и DC-SIGN способны распознавать антигены SEA, но важность этих взаимодействий in vivo до сих пор неизвестна (Meevissen et al ., 2012; Tundup et al ., 2012). Точно так же было показано, что MR связывает экскреторных / секреторных белков Trichuris muris , но не влияет на выведение паразитов, поскольку мыши MR — / — избавлялись от инфекции нормально (Deschoolmeester et al ., 2009).
Примечательно, что недавняя публикация показала роль MR против Mesocestoides corti , ленточного червя, который выделяет гликановые антигены в ЦНС, вызывая нейроцистицеркоз (Mishra et al ., 2013). In vivo , MR — / — мыши имеют повышенную выживаемость с накоплением регуляторных гранулоцитарных миелоидных клеток и сниженным количеством Т-клеток.Следовательно, патогенез нейроцистицеркоза, по-видимому, напрямую связан с иммунным ответом против паразита, индуцированным МР. Хотя эти результаты in vivo следует интерпретировать с осторожностью из-за того факта, что микроРНК (miR-511-1) кодируется в гене MR и ко-регулируется с MR, возможно, влияя на наблюдаемый фенотип (Tserel et al ., 2011).
В контексте иммунитета Th3 CLR также играют ключевую роль в стимулировании аллергических реакций.Особое значение имеют MR, DC-SIGN и Dectin-2 (Salazar et al ., 2013). В то время как MR и DC-SIGN распознают антиген Dermatophagoides pteronyssinus группы 1 (Der p 1) от клещей домашней пыли (HDM), Dectin-2 распознает как HDM ( D. farinae и D. pteronyssinus ), так и плесень. ( Aspergillus fumigatus ) экстракты. Было обнаружено, что дектин-2 запускает генерацию цистеиниловых лейкотриенов (cys-LT), которые опосредуют воспаление легких. В отсутствие LTC 4 -синтазы (критический фермент в генерации cys-LT) или рецептора cys-LT воспаление легких уменьшалось (Barret et al ., 2009).
Protozoa
Несколько CLR были вовлечены в распознавание видов Leishmania , но их роль только начинает пониматься. Молекулы DC-SIGN и L-SIGN (близкий гомолог DC-SIGN, который также распознает высокоманнозные гликаны), как было показано, распознают Leishmania , но рецепторы различаются по своей способности взаимодействовать с этими организмами, в зависимости от видов и стадии созревания паразитов (Caparros et al ., 2005). Было показано, что Dectin-1 и MR имеют решающее значение для «убивающей» реакции против L. infantum , индуцируя ROS в макрофагах и , вызывая Syk-связанную секрецию IL-1β (Lefevre et al ., 2013). ). SIGNR3, с другой стороны, обеспечивает выживаемость паразитов как in vivo , так и in vitro , посредством ингибирования оси LTB4 / IL-1β (Lefevre et al ., 2013). Поскольку известно, что LTB4 играет решающую роль в механизмах, ответственных за уничтожение Leishmania , в частности, за счет активации IL-1β; Было высказано предположение, что повышенный уровень LTB4 приносит пользу хозяину, тогда как сниженный уровень LTB4 приносит пользу патогену.Эти исследования подчеркивают различные, но важные роли CLR в патогенезе Leishmania .
CARD9 недавно был связан с церебральной малярией (ЦМ), где его экспрессия была усилена в мышиной модели, индуцированной Plasmodium Berghei (Hafalla et al ., 2012). Однако мыши CARD9 — / — не были защищены от инфекции, что позволяет предположить, что CM развивается независимо от CARD9, несмотря на его активацию во время болезни. Напротив, поразительная защита от CM наблюдалась, когда DNGR ‐ 1 + DC были истощены.Защита была связана с уменьшением количества клеток CD8 + , уменьшением количества паразитов в головном мозге и снижением уровней IFN-γ (Piva et al ., 2012). Совсем недавно исследование с использованием мышей DCIR — / — также продемонстрировало значительную выживаемость по сравнению с контролем дикого типа. Защита была связана с уменьшением количества клеток CD8 + и уменьшением воспаления мозга, подчеркивая функции активации некоторых ингибирующих рецепторов (Redelinghuys and Brown, 2011; Maglinao et al ., 2013). Хотя конкретный механизм, лежащий в основе этой защиты, неизвестен, ясно, что CLR играют критическую роль в CM.
Грибы
Врожденные и адаптивные иммунные ответы на грибки в основном опосредуются CLR, причем Dectin-1 лучше всего охарактеризован в контексте грибковых инфекций (Hardison and Brown, 2012). При грибковых инфекциях врожденный и адаптивный иммунные ответы в первую очередь регулируются CLR. В связи с быстрым ростом достижений, достигнутых в этой области, мы обсудим некоторые из недавних исследований, в которых анализируются CLR-опосредованные механизмы, участвующие в противогрибковом иммунитете.Дектин-1 распознает β-глюкан и индуцирует множество клеточных функций через свой цитоплазматический сигнальный домен и необходим для защитного иммунного ответа на Candida albicans и другие грибы у мышей и людей. Недавно мы обнаружили, что потребность в Dectin-1 в контроле C. albicans зависит от штамма, поскольку различные штаммы C. albicans имеют вариации в составе и природе своих клеточных стенок, которые проявляются только во время заражения . in vivo (Маракалала и др. ., 2013). Опосредованная Dectin-1 защита от инфекций C. albicans также недавно была приписана продукции IFN типа I инфильтрирующими почками DC, ответ, который потребовал Syk, CARD9 и IRF5 (del Fresno et al ., 2013) . Другие сигнальные CLR, которые играют важную роль, включают Dectin-2, Mincle, DC-SIGN и MR (Hardison and Brown, 2012).
Влияние грибковой микробиоты на иммунную регуляцию — еще одна область недавнего прогресса. Было обнаружено, что кишечная микробиота млекопитающих включает множество видов грибов, при этом идентифицировано более 100 известных и 100 новых видов грибов (Илиев и др. ., 2012). Было показано, что мыши, лишенные Dectin-1, имеют повышенную восприимчивость к колиту, вызванному декстрансульфатом натрия (DSS), когда в их желудочно-кишечном тракте присутствуют определенные виды грибов. Более того, был идентифицирован полиморфизм Dectin-1, связанный с пациентами с тяжелым язвенным колитом (Iliev et al ., 2012). Точно так же было показано, что распознавание грибов SIGNR3 влияет на иммунную регуляцию в кишечнике, поскольку у мышей SIGNR3 — / — наблюдается обострение вызванного DSS колита по сравнению с контрольной группой дикого типа (Eriksson et al ., 2013). Вместе эти исследования подчеркивают важность распознавания грибов CLR и роль, которую это играет в поддержании иммунного гомеостаза кишечника и контроле над заболеванием.
Как мы уже обсуждали, CLR важны для инициации врожденного иммунитета и связывают распознавание патогенов с развитием адаптивного иммунитета. Совсем недавно возникла концепция врожденной иммунной памяти или «тренированного иммунитета», которая бросила вызов традиционным парадигмам адаптивной памяти, опосредованной Т- и В-клетками.По сути, тренированный иммунитет индуцируется после первичной инфекции или вакцинации, обеспечивает защиту независимо от Т- или В-клеток, опосредованную клетками врожденного иммунитета, такими как NK-клетки и моноциты / макрофаги, и повышает сопротивляемость инфекции теми же или другими патогенами (Netea et al ., 2011; Netea, 2013). Тренированный иммунитет можно отличить от иммунного прайминга благодаря тому факту, что после выздоровления от инфекции врожденные иммунные ответы не возвращаются к устойчивому уровню.Это связано с эпигенетическим перепрограммированием клеток врожденного иммунитета, а не с кратковременным изменением состояния, наблюдаемым при иммунном праймировании. Анализ механизмов, задействованных в тренированном иммунитете, обеспечивает новый захватывающий подход к защите от инфекций. Роль CLR в тренированном иммунитете была недавно описана, где перепрограммирование моноцитов с помощью передачи сигналов Dectin-1 / Raf-1 предотвращало заражение C. albicans и другими организмами (Quintin et al ., 2012). Таким образом, понимание тренированного иммунитета, вызванного CLR, может лечь в основу новых стратегий иммунотерапии и вакцинации.
Наконец, хорошо известно, что сотрудничество между CLR и TLR инициирует оптимальные противогрибковые реакции. Фактически, совместные ответы, индуцированные Dectin-1 и TLR2, были одними из первых когда-либо описанных совместных ответов PRR (Hardison and Brown, 2012). Более свежий пример важности такого сотрудничества — признание Fonsecaea pedrosoi . Было показано, что этот организм, вызывающий хромобластомикозы, распознается CLR, но не TLR, что приводит к дефектным воспалительным ответам и восприимчивости к инфекции.Удивительно, но экзогенное введение агонистов TLR восстановило защитные воспалительные реакции и привело к исчезновению инфекции in vivo (Sousa et al ., 2011). Этот подход сейчас тестируется на людях (G.D.B., неопубликованные данные).
Полногеномное ассоциативное исследование устойчивости к инфекции Mycobacterium tuberculosis выявляет локус 10q26.2 в трех разных популяциях
% PDF-1.6 % 1 0 объект > поток DOI: 10.1371 / journal.pgen.1009392
RQd4Nra4; M + 8 Բ 2 WB /, N’x2qZI5 | [, + z_hb1 ^^ lcLX & g7beE1.-avGwf «= jO / {z ַ oȏm6skт0vuG & ‘Nͽnʩ խ آ Gi> dzʺ6gw’o-! T taW
Мыс Горн — WorldAtlas
Мыс можно определить как большой приподнятый участок суши, который простирается в океан, реку или озеро. Эти узкие прибрежные формы рельефа образованы ледниковым воздействием, эрозией или движением океанских течений. В течение нескольких лет мысы использовались моряками как важные навигационные ориентиры.
Где находится мыс Горн? Карта Огненной Земли и мыса Горн.Расположенный на острове Хорнос, мыс Горн является самым южным крутым скалистым мысом архипелага Огненная Земля в южной части Чили. Мыс Горн отмечает место встречи Тихого и Атлантического океанов, а также самый северный край пролива Дрейка.
Мыс Горн является частью коммуны Кабо-де-Орнос и входит в состав региона Магалланес-и-Антарктика-Чили.Мыс расположен в национальном парке Кабо-де-Орнос, который также является самым южным национальным парком в мире.
КлиматИз-за своего южного расположения в районе мыса Горн круглый год ветрено и прохладно.Зарегистрированы ветры со средней скоростью 30 км / час и шквалы со скоростью 100 км / час. Хотя март — самый влажный месяц с большим количеством осадков, количество осадков остается высоким в течение года.
Краткая история Мыс Горн, Огненная Земля, Чили.Мыс Горн был открыт голландским мореплавателем Виллемом Схоутеном в 1616 году.Он и его команда также первыми обогнули мыс. Схоутен назвал мыс «Kaap Hoorn» в честь голландского города Хорн. В течение нескольких лет мыс служил важной частью маршрута Клипера для судов, следующих из Европы в Восточную Азию или с одного американского побережья на другое. Однако этот навигационный маршрут вокруг мыса Горн был чрезвычайно опасным из-за наличия нескольких айсбергов, сильных ветров и течений, а также гигантских волн. Открытие Панамского канала в 1914 году значительно уменьшило потребность парусных судов в обходе мыса Горн.
Диптарка Гош 29 марта 2021 года в Landformsлектинов С-типа, грибов и ответов Th27
Резюме
Клетки Th27 представляют собой недавно обнаруженную субпопуляцию Т-хелперных клеток, характеризующихся высвобождением IL-17, и считаются важными для мобилизации иммунных ответов против микробных патогенов но которые также способствуют развитию аутоиммунных заболеваний.Идентификация рецепторов лектина С-типа, которые способны регулировать баланс между ответами Th2 и Th27, представляет особый интерес в последнее время, который они частично контролируют посредством высвобождения цитокинов, индуцирующих Th27. Многие из этих рецепторов распознают грибы и другие патогены и играют ключевую роль в развитии защитного антимикробного иммунитета. Здесь мы рассмотрим лектины С-типа, которые связаны с ответами типа Th27, и кратко рассмотрим роль ответов Th27 в противогрибковом иммунитете мышей и человека.
Ключевые слова: Th27, лектин С-типа, Syk-киназа, грибы, микобактерии
1. Введение
Эффекторные Т-хелперные клеткипервоначально были классифицированы как клетки Th2 или Th3, характеризующиеся в основном продуцированием IFNγ или IL-4. , соответственно. Совсем недавно была идентифицирована третья подгруппа эффекторных клеток, клетки Th27, которые характеризуются продуцированием IL-17. Индукция дифференцировки клеток Th27 была областью значительного внимания, и теперь понятно, что в развитии этих клеток участвуют цитокины, включая TGF-β, IL-1β, IL-6, IL-21 и IL-23, и факторы транскрипции, включая связанные с рецепторами ретиноевой кислоты орфанные ядерные рецепторы (ROR) α и γt [1].Однако фактическое участие цитокинов, таких как IL-23 и TGF-β, в дифференцировке клеток Th27 было спорным, как и родство между клетками Th27 человека и мыши, хотя многие из этих противоречий начинают разрешаться.
Помимо IL-17A и IL-17F, клетки Th27 продуцируют множество других цитокинов, таких как, например, IL-21 и IL-22, каждый из которых способен вызывать различные воспалительные и антимикробные реакции. в других типах клеток, включая миелоидные и эпителиальные клетки [1].Следовательно, ответы Th27 участвуют в управлении защитными иммунными ответами против множества микробов, включая несколько бактериальных и грибковых патогенов [2]. Однако воспалительные эффекты этих ответов также участвуют в развитии аутоиммунных заболеваний, таких как экспериментальный аутоиммунный энцефаломиелит [1]. Таким образом, защитные антимикробные эффекты иммунитета Th27 также могут вносить вклад в патологию, что, возможно, объясняет некоторые противоречия, связанные с ролью этих ответов в мышиных моделях грибковой инфекции (обсуждаются позже).
Другой интересной областью была идентификация рецепторов распознавания образов (PRR) и внутриклеточных сигнальных путей в антигенпрезентирующих клетках (APC), которые управляют развитием иммунитета Th27. Одна группа PRR, которые, по-видимому, играют все более важную роль в этих типах ответов, — это рецепторы лектина C-типа (CLR), и особенно те, которые передают сигнал через путь Syk / CARD9. Как мы увидим, многие из этих рецепторов участвуют в распознавании грибов и бактерий и способны напрямую стимулировать развитие адаптивных иммунных ответов.Таким образом, эти рецепторы действуют как архетипические PRR с функциями, аналогичными функциям семейства Toll-подобных рецепторов (TLR). В этом обзоре мы рассмотрим каждый из CLR, которые участвуют в иммунитете Th27, а также кратко обсудим, что известно о роли этого типа адаптивного ответа в противогрибковом иммунитете.
2. Лектины C-типа и ответы Th27
Лектин-подобные рецепторы C-типа (CLR) представляют собой разнообразное семейство белков, которые характеризуются наличием одного или нескольких лектин-подобных доменов C-типа (CTLD). .CTLD обычно участвует в распознавании лиганда и состоит из отдельной белковой складки, которая создается дисульфидными мостиками между консервативными остатками цистеина [3]. CLR обнаруживаются как растворимые, так и связанные с мембраной белки, и, хотя первоначально они определялись как белки, связывающие углеводы, теперь известно, что они способны распознавать широкий спектр других экзогенных патоген-ассоциированных молекулярных паттернов (PAMP) и эндогенных лигандов, включая белки и липиды. . Некоторые из мембраносвязанных рецепторов способны запускать внутриклеточную передачу сигналов; либо непосредственно через интегральные цитоплазматические сигнальные мотивы, либо опосредованно, через ассоциацию с сигнальными адапторными молекулами, такими как цепь Fcγ (см.[4] для обзора). Внутриклеточные сигнальные пути, индуцированные этими рецепторами, могут либо подавлять клеточную функцию (что обычно происходит с рецепторами, содержащими иммунорецепторные ингибирующие мотивы на основе тирозина или ITIM), либо эти рецепторы могут запускать клеточную активацию (через иммунорецепторные мотивы активации на основе тирозина или ITAMs), что приводит к стимуляции различных клеточных ответов, включая индукцию экспрессии генов. Основываясь на их филогении и доменной организации, CLR были разделены на 17 семейств [5], но представляют интерес члены рецепторов группы II, V и VI, которые участвуют в ответах типа Th27, включая Dectin-1, Dectin-2. , Mincle, DC-SIGN, CLEC-1 и CD161 (краткое изложение ролей и лигандов этих различных рецепторов показано на и).Большинство этих рецепторов также вовлечены в противомикробный иммунитет.
Мультяшное изображение ролей CLR во время разработки ответов Th27.
Таблица 1
Лектиновые рецепторы С-типа (CLR), их лиганды и их роль в развитии ответов Th27.
| CLR | Лиганды | Распознанные патогены | Роль в ответах клеток Th27 |
|---|---|---|---|
| Дектин-1 | Экзогенный : β1,3-глюкан | 09 Грибы Aspergillus , Pneumocystis , Coccidioides , Saccharomyces | Способствует ответам Th27 на грибки и микобактерии |
| Эндогенный : лиганд Т-клеток? | Бактерии : микобактерии | ||
| Дектин-2 | Экзогенные : структуры с высоким содержанием маннозы | Грибки : Cryptococcus , 9000sporphy Candida , Paracoccidioides, Histoplasma | Способствует ответам Th27 на грибки |
| Эндогенный : лиганд Т-клеток? | Бактерии : микобактерии | ||
| Рецептор маннозы | Экзогенный : терминальная манноза, фукоза, N-ацетилглюкозамин, сульфатированные сахара. , Pneumocystis | Неясно: подавляет ответы Th27 на микобактерии, но вызывает эти ответы на Candida . | |
| Эндогенный : различные, например гликопротеиновые рецепторы, гормоны, лизосомные ферменты | Бактерии : микобактерии, клебсиелла, стрептококки | ||
| Вирус : денге, ВИЧ | |||
| 2 | |||
| Mincle | Экзогенный : α-маннан, микобактериальный пуповинный фактор | Грибы : Candida , Saccharomyces , Malassezia | mycobacterium |
| Эндогенный : SAP130 | |||
| DC-SIGN | Экзогенный : высокоманнозные и фукозилированные структуры. | Fungi : Candida , Chrysosporium, Aspergillus | Неясно: но может подавлять ответы Th27. |
| Эндогенный : различные, например молекула межклеточной адгезии (ICAM) -2 и 3, Lewis-x | Бактерии : микобактерии | ||
| Вирус : ВИЧ | |||
| CLEC-1 2 | Экзогенный 😕 | ? | Неясно: а может подавить ответы Th27? |
| Эндогенный : Т-клеточный лиганд? | |||
| CD161 | Экзогенный 😕 | ? | Маркер Th27 |
| Эндогенный : PILAR, CLEC2D | Т-клеточная пролиферация? | ||
| Миграция клеток? | |||
| Регулирование производства цитокинов? | |||
2.1. Дектин-1 (Clec7a)
Дектин-1 представляет собой трансмембранный белок типа II, который экспрессируется преимущественно миелоидными клетками (моноцитами / макрофагами, дендритными клетками и нейтрофилами), хотя рецептор экспрессируется на некоторых других типах клеток, включая различные популяции лимфоцитов [6]. Наивысшие уровни экспрессии Dectin-1 наблюдаются в воспалительных клетках и в клетках на портах проникновения патогенов, и уровни этого рецептора могут значительно модулироваться клеточным созреванием, цитокинами и другими модификаторами биологического ответа [6].Примечательно, что экспрессия Dectin-1 может быть индуцирована на других типах клеток, включая эпителий слизистой оболочки [7].
Dectin-1 состоит из одного внеклеточного CTLD, связанного ножкой и трансмембранной областью с цитоплазматическим хвостом, содержащим ITAM-подобный мотив (также называемый hemITAM;) [6]. Рецептор существует в виде двух основных изоформ, генерируемых альтернативным сплайсингом, которые отличаются включением внеклеточной области ножки и обладают небольшими функциональными различиями [6]. Дектин-1 специфически распознает β1,3-связанные глюканы, углеводы, обнаруженные в клеточных стенках грибов, растений и некоторых бактерий.Dectin-1 распознает несколько видов грибов, включая ряд патогенов человека, таких как Candida , Aspergillus , Pneumocystis и Coccidioides , и играет важную роль в защите хозяина от этих патогенов [6,8,9] . Дектин-1 также может распознавать микобактерии и эндогенную молекулу на Т-клетках, но идентичность задействованных лигандов неизвестна [6].
Карикатурное изображение структур и известных сигнальных путей, которые используются различными CLR, участвующими в ответах Th27.
Связывание лиганда с Dectin-1 может вызывать различные клеточные реакции, включая поглощение лиганда за счет эндоцитоза и фагоцитоза, клеточное созревание, респираторный взрыв, продукцию метаболитов арахадоновой кислоты и продукцию многочисленных цитокинов и хемокинов, включая IL-2 , IL-10, CXCL2, TNF, IL-1β, IL-6 и IL-23 [6]. Дектин-1 может также взаимодействовать с MyD88-сопряженными Toll-подобными рецепторами, синергически индуцируя выработку цитокинов, включая IL-6 и IL-23, но эти взаимодействия также приводят к понижающей регуляции IL-12, что, вероятно, способствует развитие ответов Th27 (см. ниже) [10,11].Передача сигналов от Dectin-1 опосредуется цитоплазматическим ITAM-подобным мотивом и включает несколько нисходящих путей, включая те, которые опосредуются через Syk / CARD9 и Raf-1 киназу [6].
Дектин-1 также способен направлять развитие адаптивного иммунитета. Было показано, что стимуляция Dectin-1 на антигенпрезентирующих клетках (APC) с использованием высокоочищенных β-глюканов индуцирует дифференцировку Th27 и Th2 CD4 + T-клеток и стимулирует развитие антител и CD8 + T-клеток. ответы [12–14].Как Dectin-1 на самом деле управляет развитием Th27-ответов, все еще неясно, хотя для этого необходимы сигнальные пути киназы Syk / CARD9 и Raf-1, фосфолипаза C-γ2, и, вероятно, это связано со способностью Dectin-1 стимулировать (IL-1β, IL-6 и IL-23) и ингибировать (IL-12) выработку цитокинов, важных для формирования этих ответов [11,14,15]. Также было показано, что стимуляция Dectin-1 на APC и генерация IL-23 способна управлять преобразованием выбранных популяций клеток T reg в T-клетки, продуцирующие IL-17 [16].Совсем недавно прямая стимуляция Dectin-1 на γδ Т-клетках, которые, как полагают, вносят вклад в ранние врожденные ответы, индуцировала продукцию IL-17 из этих клеток; ответ, который может быть значительно усилен в присутствии IL-23 [17].
Несколько исследований подтвердили роль Dectin-1 в управлении ответами IL-17 во время грибковой инфекции. У людей был идентифицирован полиморфизм Dectin-1, который вводит преждевременный стоп-кодон (Tyr238X), который предотвращает экспрессию этого рецептора на поверхности клетки [9].Лица, гомозиготные по этому полиморфизму, восприимчивы к грибковым инфекциям слизистых и кожных покровов и имеют дефектную продукцию цитокинов, включая IL-17, в ответ на грибки [9]. Совсем недавно этот полиморфизм и его влияние на продукцию IL-17 были связаны с восприимчивостью к инвазивному аспергиллезу у пациентов с трансплантацией стволовых клеток [18], а также со снижением частоты реакции трансплантат против хозяина, которая была связана с Candida колонизация у этих пациентов [19].Интересно, что в моделях на мышах Dectin-1, по-видимому, не играет важной роли в формировании иммунитета Th27 в ответ на C. albicans . [12], однако этот рецептор участвовал в выработке защитных ответов IL-17 во время экспериментальной легочной инфекции A. fumigatus [8].
Помимо грибов, Dectin-1 участвует в развитии адаптивных ответов на Mycobacterium tuberculosis . Несколько исследований показали, что Dectin-1 участвует в поглощении микобактериями и индукции цитокинов, таких как IL-12, IL-23, IL-1β и IL-6, в ответ на эти организмы [6,20].Кроме того, было показано, что распознавание микобактерий Dectin-1 на APC человека in vitro способствует образованию Th2 и Th27 CD4 + лимфоцитов, и рецептор также может участвовать в опосредованной микобактерией стимуляции IL-17 из γδ Т-клетки [17,20,21].
2.2. Дектин-2 (Clec-4n)
Дектин-2 представляет собой трансмембранный рецептор типа II, экспрессируемый преимущественно на тканевых макрофагах, DC и воспалительных моноцитах [22]. Рецептор обладает классическим сахаросвязывающим CTLD, который распознает структуры с высоким содержанием маннозы зависимым от Ca 2+ образом, посредством чего он распознает множество патогенов, включая дефицитные по капсуле Cryptococcus neoformans , Candida albicans , Saccharomyces cerevisiae , м.tuberculosis , Microsporum audounii , Trichophyton rubrum , Paracoccidioides brasiliensis и Histoplasma capsulatum [23,24]. Дектин-2 также распознает аллергены клещей и грибов домашней пыли и играет роль в устойчивости к УФ-излучению за счет распознавания неидентифицированного эндогенного лиганда на CD4 + CD25 + Т-клетках [25, 26]. Хотя рецептор имеет короткий цитоплазматический хвост, в котором отсутствуют традиционные сигнальные мотивы, Dectin-2 связывается с ITAM-содержащим адаптером FcRγ и может запускать внутриклеточную передачу сигналов через путь Syk-CARD9, вызывая различные клеточные ответы, включая продукцию экозаноидов, цитокины и хемокины () [24,26–30].
Два недавних исследования, сфокусированных на характеристике роли Dectin-2 в ответе хозяина на C. albicans , предоставили первые доказательства роли этого рецептора в индукции ответов Th27. В этих исследованиях было показано, что дектин-2 способствует индукции цитокинов, включая TNF, IL-6, IL-1β и IL-23, после стимуляции DC частицами грибов. Важно отметить, что передача сигналов Dectin-2 через путь Syk / CARD9, как было показано, играет существенную роль в развитии ответов Th27 и Th2 на C.albicans у мышей, хотя последний ответ также требовал Dectin-1 [28,29]. Используя мышей с дефицитом гена, Saijo et al . дополнительно смогли показать, что Dectin-2, который распознает грибковые α-маннаны, необходим для устойчивости к инфекции C. albicans [28]. Основываясь на этих результатах, Robinson et al . предположили, что Dectin-2 и Dectin-1 объясняют почти всю Syk / CARD9-зависимую передачу сигналов, индуцируемую в ответ на эти грибковые патогены [29].
2.3. Рецептор маннозы
Рецептор маннозы макрофагов (MR; CD206) представляет собой трансмембранный белок типа I, который имеет восемь внеклеточных CTLD и короткий цитоплазматический хвост, в котором отсутствуют классические сигнальные мотивы (). Хотя большая часть MR находится внутри клетки, внутри эндоцитарного пути, растворимая форма рецептора также попадает в сыворотку. MR экспрессируется макрофагами, некоторыми DC, а также множеством других клеток и тканей, включая печеночный и лимфатический эндотелий [31,32].MR может связывать терминальную маннозу, фукозу или N-ацетилглюкозамин и, следовательно, распознает широкий спектр эндогенных и экзогенных лигандов, включая несколько бактериальных, вирусных и грибковых патогенов (таких как C. neoformans , C. albicans и Pneumocystis). carinii ) [32].
Было показано, что MR вызывает множество клеточных ответов, но молекулярные механизмы, ответственные за передачу внутриклеточных сигналов от этого рецептора, неясны.Было показано, что распознавание грибов этим рецептором, которое может происходить только в фагосоме после поглощения грибами, способствует выработке ряда цитокинов, таких как TNF, GM-CSF, IL-12, IL-8, IL-6. , и IL-1β, хотя есть также свидетельства того, что MR может ингибировать продукцию определенных цитокинов, включая TNF [32–34]. Хотя MR играет очевидную роль в гомеостазе, его роль в антимикробном иммунитете все еще неясна, поскольку мыши с дефицитом MR не демонстрируют значительных изменений иммунитета к C.albicans или P. carinii [35–37]. Однако было обнаружено, что эти животные более восприимчивы к заражению C. neoformans , что является результатом неправильной индукции защитных ответов Т-клеток CD4 + [38].
Использование анализов in vitro с человеческими PBMC, Netea et al . недавно продемонстрировали, что стимуляция MR с помощью Candida или очищенного маннана Candida способна индуцировать значительные уровни IL-17 [39].Этот ответ не был прямой митогенной стимуляцией Т-клеток, поскольку он требовал APC, и как Dectin-1, так и TLR2 были способны усиливать эти ответы. Этот ответ был специфическим для Candida , так как маннан, выделенный из S. cerevisiae , не имел никакого эффекта [39]. Однако, в отличие от этих результатов, другое исследование продемонстрировало, что стимуляция MR может подавлять ответы Th27, индуцированные микобактериями [20]. Понимание сигнальных механизмов, используемых MR, и эффектов сотрудничества с разными PRR, несомненно, являются проблемами, которые необходимо решить, чтобы полностью понять роль этого рецептора.
2.4. Mincle (Clec4e)
Лектин C-типа, индуцируемый макрофагами (Mincle), представляет собой трансмембранный белок типа II, который в первую очередь экспрессируется активированными макрофагами и, вероятно, также DCs [40,41]. Подобно Dectin-2, Mincle обладает одним внеклеточным CTLD, коротким цитоплазматическим хвостом и ассоциируется с адаптером FcRγ, чтобы запускать внутриклеточную передачу сигналов через путь Syk / CARD9 [42] (). Mincle распознает множество эндогенных и экзогенных лигандов, таких как некротические клетки, микобактерии и некоторые грибы, включая Candida , Saccharomyces и Malassezia (хотя рецептор может предпочтительно распознавать последний вид грибов) [42–44].Многие из лигандов, участвующих в этих взаимодействиях, были идентифицированы и включают грибковый α-маннан, микобактериальный пуповинный фактор (димиколат трегалозы, а также синтетический аналог трегалозы-дибегенат) и малый ядерный рибонуклеопротеин SAP130 [42,44–46]. Было показано, что при распознавании этих лигандов Mincle вызывает множество клеточных ответов, включая индукцию цитокинов, таких как TNF, MIP-2, KC, IL-10 и IL-6. Мыши с нокаутом Mincle проявляют повышенную восприимчивость к инфекциям, вызванным Candida , а блокирование функции Mincle с помощью антител in vivo снижает рекрутирование нейтрофилов и продукцию воспалительных цитокинов в ответ на некротическую гибель клеток [42,43].
Здесь важна способность Mincle опосредовать иммунные ответы на трегалозодимиколат (TDM) и трегалозодибегенат (TDB). TDM, который обладает мощной воспалительной активностью и считается ключевым фактором патогенеза при туберкулезе, и его менее токсичный аналог TDB оказались полезными адъювантами для микобактериальных субъединичных вакцин в стимулировании развития защитных ответов Th2 и Th27 [41 , 47]. Недавние данные показали, что Mincle опосредует все ответы на TDM и TDB, при этом мыши с дефицитом Mincle (или цепи FcRγ) теряют способность индуцировать ответы Th2 и Th27 после иммунизации TDB / антигеном [45,46].Таким образом, как и другие лектины С-типа, связанные с Syk, описанные выше, Mincle способен направлять развитие ответов Th27, и, хотя пока это показано только для микобактерий, этот рецептор, вероятно, также вносит вклад в развитие этих ответов во время грибковой инфекции. .
2,5. DC-SIGN
DC-SIGN человека (CD209) представляет собой трансмембранный белок типа II, обладающий одним внеклеточным CTLD и цитоплазматическим хвостом, содержащим мотивы интернализации (). DC-SIGN экспрессируется в виде тетрамера из-за взаимодействий между внеклеточными стеблевыми областями мономеров, и рецептор экспрессируется в основном незрелыми DC, но он также обнаруживается на отдельных макрофагах и эндотелиальных клетках [48-50].Мыши экспрессируют восемь ортологов этого рецептора, которые немного различаются по своей структуре и профилям экспрессии [51,52]. DC-SIGN распознает углеводы, включая высокоманнозные и фукозилированные структуры, что позволяет ему распознавать широкий спектр патогенов, включая микобактерии и несколько грибковых патогенов, таких как видов Candida , конидии A. fumigatus , Chrysosporium tropicum , и, возможно, также C. neoformans [51,53–55].
Хотя точные механизмы все еще неясны, DC-SIGN способен индуцировать внутриклеточную передачу сигналов, приводящую к клеточным ответам, таким как фагоцитоз [55].Передача сигналов DC-SIGN частично опосредуется через путь киназы Raf-1 и может модулировать продукцию цитокинов, индуцированную другими PRR, включая TLR, хотя рецептор, по-видимому, не способен напрямую индуцировать продукцию цитокинов [56 ]. Передача сигналов, индуцированная DC-SIGN, зависит от природы углеводного лиганда, что приводит к стимуляции или репрессии ряда цитокинов, включая IL-10, IL-12 и IL-6 [56,57]. Действительно, недавнее исследование продемонстрировало, что стимуляция DC-SIGN с использованием определенных лигандов может ингибировать продукцию цитокинов, включая IL-1β, IL-23 и TNF, которые индуцируются M.tuberculosis или бета-глюкан в ДК человека, и это привело к репрессии ответов Th27 [20]. Таким образом, хотя эти результаты продемонстрированы только в одном исследовании на сегодняшний день, они предполагают, что передача сигналов от DC-SIGN может модулировать развитие ответов Th27.
2.6. CLEC-1 (CLEC-1A)
CLEC-1 является относительно плохо охарактеризованным членом кластера Dectin-1 лектинов C-типа и экспрессируется эндотелиальными клетками, DC и миелоидными клетками других тканей, но не моноцитами, гранулоциты, В-, Т- или NK-клетки периферической крови [58,59].Этот трансмембранный рецептор типа II структурно подобен Dectin-1, хотя он не содержит каких-либо канонических сигнальных мотивов и может ассоциироваться с адаптером для экспрессии на поверхности клетки [58]. Однако цитоплазматический хвост действительно содержит единственный остаток тирозина, который может индуцировать внутриклеточную передачу сигналов (). Было обнаружено, что экспрессия CLEC-1 подавляется воспалительными стимулами, такими как LPS или IFNγ, но усиливается иммуносупрессивными цитокинами, такими как IL-10 и TGF-β [60].Интересно, что на моделях грызунов было обнаружено, что CLEC-1 активируется в переносимых аллотрансплантатах, и это было связано с низкими уровнями IL-17 и присутствием CD4 + CD25 + T reg клеток в месте трансплантата. . Анализы In vitro продемонстрировали, что экспрессия CLEC-1 увеличивалась на эндотелиальных клетках при контакте с клетками CD4 + CD25 + T reg , и что потеря экспрессии CLEC-1 из DC (с использованием siRNA) усиливала развитие клеток Th27 при снижении экспрессии Foxp3 в MLR [60].В целом эти результаты предполагают, что CLEC-1 играет роль в регуляции развития клеток Th27, но необходимы дополнительные данные, чтобы полностью понять роль и лежащие в основе механизмы, используемые этим рецептором.
2.7. CD161
CD161 (NKRP1a) — трансмембранный рецептор типа II, который состоит из одного внеклеточного CTLD и цитоплазматического хвоста, который содержит атипичный ITIM-подобный мотив [61] (). Рецептор экспрессируется подмножествами NK, NKT и T-клеток, а также обнаружен на моноцитах и DC [61–63].Было показано, что CD161 распознает ряд лигандов, включая углеводы, лектин-подобный транскрипт-1 (LLT1, CLEC2D) и рецептор, связанный с лимфоцитами, индуцированный пролиферацией (PILAR) [64–66]. Роль CD161 все еще неясна, но было показано, что он способен регулировать цитолиз NK-клеток, трансэндотелиальную миграцию и пролиферацию незрелых тимоцитов, Т-клеток и NKT-клеток [62,64,65,67,68 ]. Рецептор также может модулировать продукцию цитокинов, включая TNF, IL-1β, IFNγ, IL-4, IL-12, в различных миелоидных и лимфоидных клетках CD161 + [62,69].Как CD161 опосредует эти эффекты, все еще в значительной степени неизвестно, хотя было показано, что его влияние на пролиферацию Т-клеток включает взаимодействие с PILAR, а передача сигналов в NK-клетках, как было показано, включает активацию кислой сфингомиелиназы [66,70].
Здесь актуальным является использование CD161 в качестве маркера человеческих Т-клеток, продуцирующих IL-17. Первоначально было продемонстрировано определение подмножеств CD4 + и CD8 + человеческих Т-клеток с различной функциональной активностью; впоследствии было показано, что CD161 имеет высокую регуляцию в Т-клетках, продуцирующих ИЛ-17, и в клетках-предшественниках, обнаруженных в пуповине. пуповина и вилочковая железа новорожденного [71,72].Это привело к предположению, что CD161 был специфическим маркером для клеток Th27, и последующие исследования показали, что этот рецептор действительно экспрессируется на всех подмножествах Т-клеток, продуцирующих IL-17 человека, включая те, которые участвуют в воспалении кишечника и в хроническом воспалении, вызванном вирусная инфекция [72–75]. Интересно, что мышиный гомолог (NK1.1) не был обнаружен на Т-клетках, продуцирующих IL-17, что внесло свой вклад, по крайней мере частично, в предположение, что мышиные клетки Th27 отличаются от клеток человека [76].
Несмотря на использование этого рецептора в качестве маркера Т-клеток Th27, его функция в отношении этих клеток все еще неясна. Стимуляция этого рецептора на клетках Th27 с помощью антител не влияла на пролиферацию или продукцию цитокинов [72]. Учитывая, что клетки Th27 могут нацеливаться на определенные ткани посредством экспрессии хемокиновых рецепторов, таких как CCR6, Cosmi et al . предположили, что CD161 может играть роль в трансэндотелиальной миграции этих клеток [72–74]. Очевидно, что еще многое предстоит узнать о функции CD161 и его участии в этих ответах.
Полногеномное ассоциативное исследование устойчивости к инфекции Mycobacterium tuberculosis идентифицирует локус 10q26.2 в трех разных популяциях
% PDF-1.7 % 1 0 объект > / Метаданные 4 0 R / Страницы 2 0 R / StructTreeRoot 3 0 R / Тип / Каталог / Средство просмотра Предпочтения 5 0 R >> эндобдж 4 0 obj > поток Microsoft® Word для Microsoft 365 Microsoft® Word для Microsoft 3652020-07-14T15: 23: 21Z2021-07-22T18: 37: 56-07: 002021-07-22T18: 37: 56-07: 00uuid: A599D2A7-E124-45DF- 8CCE-EDCE64F2E64Buuid: b8b38ab4-1dd1-11b2-0a00-aa0000000000application / pdf
Захватывающие дух виды на живописную петлю с тремя мысами
Посмотреть всю береговую линию ТилламукаПо мере того, как живописная петля продолжается от Тилламука к Оушенсайду, вы будете в восторге от завораживающих видов на океан, которые веками лишали людей дара речи.Наблюдайте, как океан встречает безмятежный залив Нетартс (остановитесь на некоторое время и отправляйтесь молчать в этом популярном месте для ловли моллюсков — просто убедитесь, что у вас есть лицензия, прежде чем отправиться в путь!), Пока вы путешествуете до мыса Лукаут, где вы находитесь в отличной позиции, чтобы увидеть вся береговая линия Тилламука. Прогуляйтесь по старовозрастному лесу и вдоль пляжа, чтобы размять ноги.
Отсюда дорога приведет вас через дюны Сандлейк в Пасифик-Сити, где скала Стог Хей выступает из глубин Тихого океана на мысе Киванда.Выпейте, любуясь великолепным видом на эту знаменитую прибрежную достопримечательность.
Легкая дневная поездка с множеством крутых остановокХотя поездка по живописной петле «Три мыса» — легкий день, по пути стоит остановиться в нескольких исторических местах и государственных парках, чтобы размять ноги. В конце концов, самые лучшие виды открываются не с пассажирского сиденья автомобиля или фургона, а после короткого перехода по лесной тропе.
В начале своего путешествия вы пройдете мимо парка Bay Ocean Park, ныне вымершего поселения, которое когда-то было «Атлантическим городом на западе».«Посмотрите, что осталось от этого района, поскольку здания были разрушены в 30-40-х годах ветром и приливами (песок, легко перемещаемый этими силами, является нестабильным фундаментом — жители и владельцы бизнеса усвоили это на собственном горьком опыте). Путешествуя на юг, вы увидите мыс Мирес с его короткими пешеходными тропами, знаменитым маяком и гигантским осьминогом — елью ситкинской породы, которая приобрела самую необычную форму! Тропа в этом районе, прямо перед тем, как вы доберетесь до парковки, приведет вас к крутому переходу к пляжу.
Еще одно место, куда стоит сходить? Пивоварня Pelican и Taproom на мысе Киванда, единственный пивной паб на побережье Северо-Запада! Вы можете насладиться обедом и выпить пинту, любуясь видом на скалу Хейстек и слушая океан.
Наконец, парк штата Боб Страуб в Пасифик-Сити предлагает красивый пляжный участок, где вы можете охотиться за ракушками и песочными долларами, греясь на солнце (или наслаждаясь небольшим моросящим дождем в Орегоне!)
Виды потрясающие в любую погодуВиды невероятные при любых погодных условиях, хотя в ясную погоду с вершины мыса Лукаут открывается самый дальний вид на побережье.Но в любое время года вы проезжаете по живописной петле «Три мыса», оно того стоит. Не забудь свой фотоаппарат! Вы не найдете более великолепного участка на побережье Орегона.
% PDF-1.4 % 520 0 объект > эндобдж xref 520 112 0000000016 00000 н. 0000003787 00000 н. 0000003875 00000 н. 0000004545 00000 н. 0000004716 00000 н. 0000004862 00000 н. 0000004919 00000 н. 0000004956 00000 н. 0000069424 00000 п. 0000069453 00000 п. 0000069626 00000 п. 0000069772 00000 п. 0000069829 00000 п. 0000070002 00000 п. 0000070148 00000 п. 0000070205 00000 п. 0000070355 00000 п. 0000070500 00000 н. 0000070557 00000 п. 0000070706 00000 п. 0000070851 00000 п. 0000070908 00000 п. 0000071058 00000 п. 0000071202 00000 п. 0000071259 00000 п. 0000071409 00000 п. 0000071553 00000 п. 0000071610 00000 п. 0000071760 00000 п. 0000071904 00000 п. 0000071961 00000 п. 0000072111 00000 п. 0000072255 00000 п. 0000072312 00000 п. 0000072461 00000 п. 0000072606 00000 п. 0000072663 00000 п. 0000072813 00000 п. 0000072957 00000 п. 0000073014 00000 п. 0000073164 00000 п. 0000073308 00000 п. 0000073365 00000 п. 0000073513 00000 п. 0000073658 00000 п. 0000073715 00000 п. 0000073863 00000 п. 0000074008 00000 п. 0000074065 00000 п. 0000074231 00000 п. 0000074385 00000 п. 0000074445 00000 п. 0000074648 00000 п. 0000075083 00000 п. 0000075551 00000 п. 0000075965 00000 п. 0000076320 00000 п. 0000077198 00000 п. 0000077696 00000 п. 0000078287 00000 п. 0000079233 00000 п. 0000079676 00000 п. 0000079905 00000 н. 0000081161 00000 п. 0000081246 00000 п. 0000081382 00000 п. 0000081575 00000 п. 0000082757 00000 п. 0000083336 00000 п. 0000083673 00000 п. 0000083881 00000 п. 0000084168 00000 п. 0000085266 00000 п. 0000085455 00000 п. 0000085869 00000 п. 0000085942 00000 п. 0000086655 00000 п. 0000086879 00000 п. 0000087320 00000 п. 0000087576 00000 п. 0000088685 00000 п. 0000089850 00000 п. 0000089987 00000 н. 00000
00000 п.